- Силовое поле используемое при построении топологии - amber99sb.
- Заряд системы -10 (обусловлен наличием 5 фосфатов в ДНК). Для нейтрализации системы внесла 10 положиетльных зарядов.
- Размер и форму ячейки : почти куб с размерами 5.014*5.007*5.268 (nm) V= 132,254 nm^3 (из файла dna_ec.gro)
- Минимизация энергии: (из файла dna_em.log)
- Алогритм минимизации энергии. integrator=l-bfgs; steepest descent algoritm for energy minimization
- Алгоритм расчёта электростатики и Ван-дер-Ваальсовых взаимодействий. coulombtype= Cut-off (для электростатики); vdw-type = Cut-off
- Утряска растворителя:
- Для биополимеров параметр, обуславливающий неподвижность биополимера. -DPOSRES
- Число шагов. 10000; total 1.0 ps.
- Длина шага. 0.001; ps
- Алгоритм расчёта электростатики и Ван-дер-Ваальсовых взаимодействий. coulombtype = pme; vdw-type = Cut-off
- Алгоритмы термостата и баростата. Tcoupl = Berendsen, Pcoupl = no
- Основной расчёт МД:
- Время моделирования 9 hours 17 minutes 41 seconds, количество процессоров 16, эффективность маштабирования.
- Длину траектории 20 ns
- Число шагов. = 10000000; total 1.0 ps.
- Длина шага. = 0.002; ps !
- Алгоритм интегратора. = md
- Алгоритм расчёта электростатики и Ван-дер-Ваальсовых взаимодействий. coulombtype= pme; vdw-type = Cut-off
- Алгоритмы термостата и баростата. Tcoupl= v-rescale; Pcoupl= Berendsen
Анализ визуального движения молекул - файл pdb
|
Модель 1 0 ns |
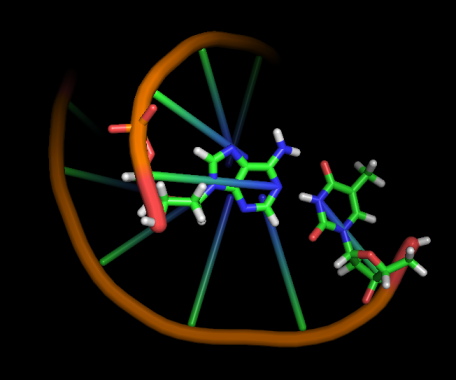
|
Модель 17 3.2 ns |
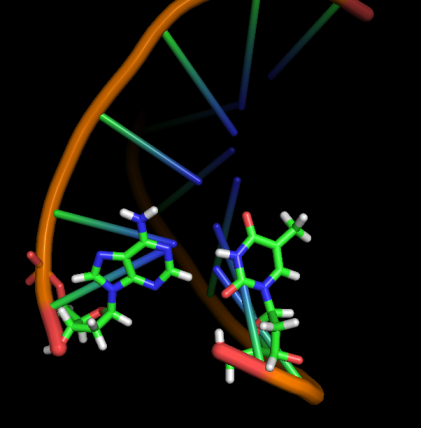
|
|
Модель 18 3.4 ns |
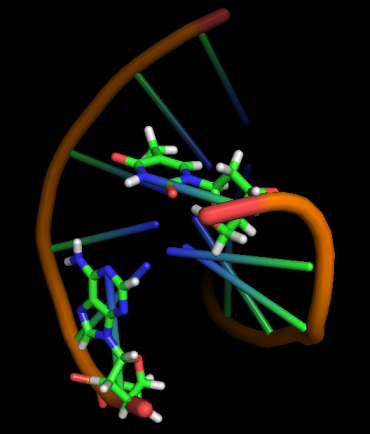
|
Модель 33 6.4 ns |
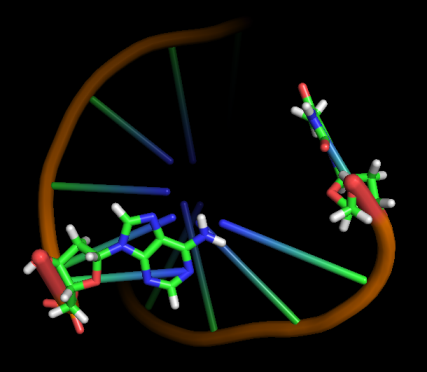
|
|
Модель 45 8.8 ns |
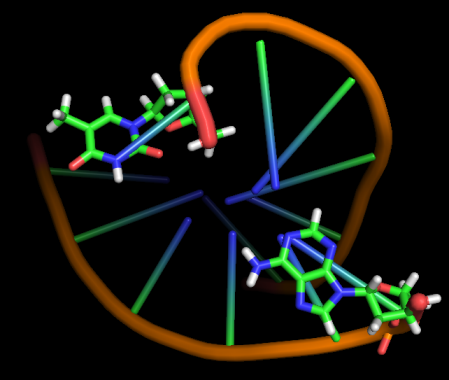
|
Модель 49 9.6 ns |
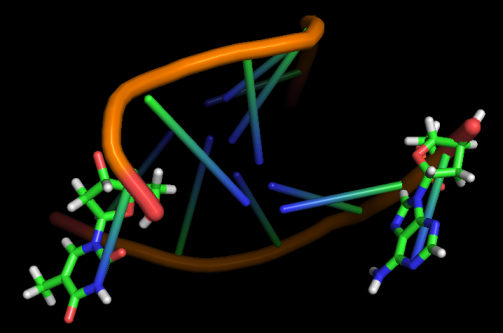
|
|
Модель 70 13.8 ns |
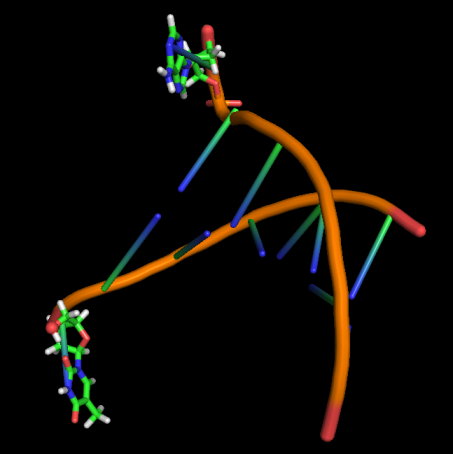
|
Модель 101 20 ns |
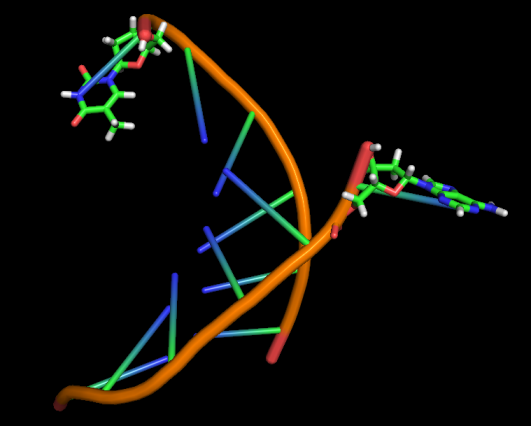
|
Первые изменения структуры, касающиеся положения относительно друг друга пары нуклеотидов GC, происходят к 17-18 модели - из одной плоскости они переходят в разные. К 33 модели цитозин из этой 1 пары начинает выворачиваться наружу, и его выворачивание заканчивается к 45 модели. С 49-50 модели начинается выворачивание и гуанина. Итого, к 70 модели происходят наибольшие изменения в структуре, приводящие к разраву водородных связей пары G-C и расхождению 1 нуклеотида в последовательности. К 20 ns происходят изменения во всей структуре, остов гнется, пары комплементарных оснований выходят из одной плоскости, цепи расходятся в районе 1 нуклеотида.
Cредне-квадратичное отклонение
Определено средне-квадратичное отклонение в ходе моделирования. RMS1- отклонение в ходе всей симуляции относительно стартовой структуры, RMS2- относительно предыдущей структуры на расстоянии 400 кадров:
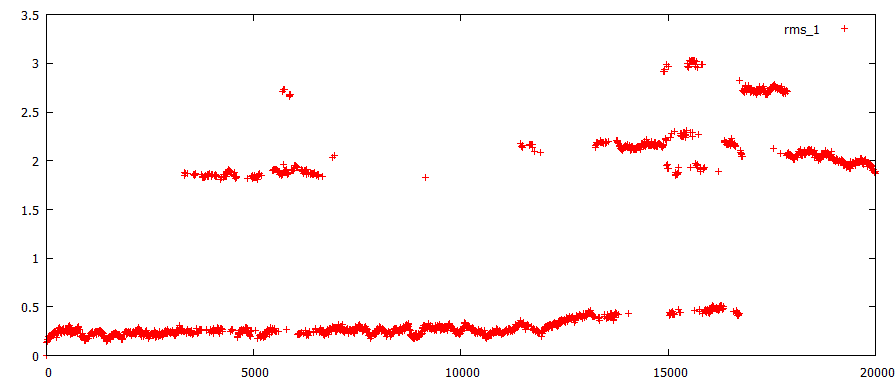
Первые явные изменения относительно начальной структуры происходят в районе 5 ns, т.е. в момент времении, соответствующий 17-33 моделям - происходят первые смещения нуклеотидов GC относительно друг-друга. Следующие значительные изменения в районе 14 ns - соответствуют модели 70, к которой 1 пара оснований выворачивается изнутри спирали ДНК наружу. Далее все структуры значительно отличаются от начальной, т.е. произошли значимые изменения в структуре исходной ДНК, которые не обращаются обратно в исходное положение.
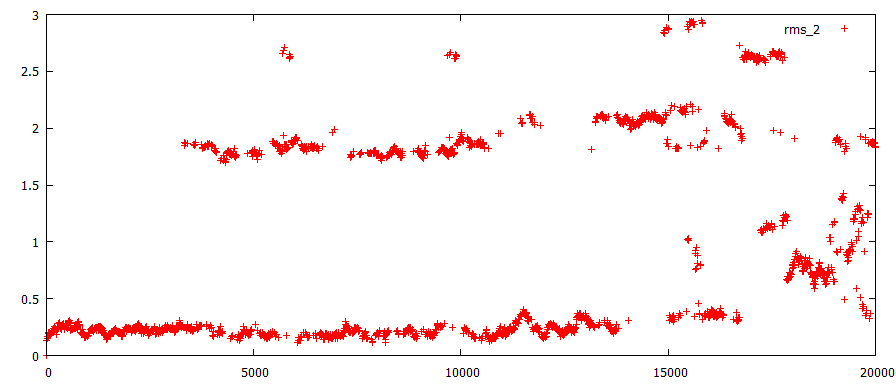
Если смотреть изменение структур огтносительно предыдущей (за 400 кадров до), то видно, что самые значительные изменения начинаются с 3 ns и постоянно продолжаются до 16 ns. Далее изменения становятся меньше, а вообще говоря, появляется разброс в их значении, т.е. струтура становится нестабильна, но происходят меньшие изменения. Это соответствует моделям после 70, в которых основные изменения - расхождение первой пары - уже произошли, а дальше не намного меняется лишь остов. Судя по тому, что значение отклонения не снижается до какого-то фонового значения, можно предположить, что и дальше должено происходить расплетение цепей.
Зависимость гидрофобных и гидрофильных поверхностей
Построена зависимость гидрофобных и гидрофильных поверхностей, доступных растворителю в любой момент времени
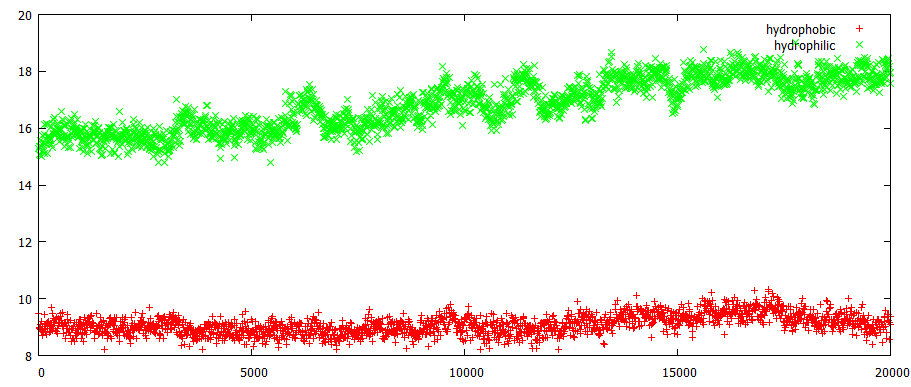
Гидрофобных поверхностей остается примерно столько же на всем учатстке, а вот для гидрофильных наблюдается рост. Возможно, это связано с тем, что гетероциклические основания выводятся изнутри спирали, происходит разрушение между ними водородных связей, и теперь растворителю становится доступной большая площадь. Это предположение подтверждается началом роста гидрофильной поверхности: где-то около 6 ns - что соответствует модели 30-33, начиная с которой, происходит выворачивание основания наружу.
Расчет количества образуемых водородных связей
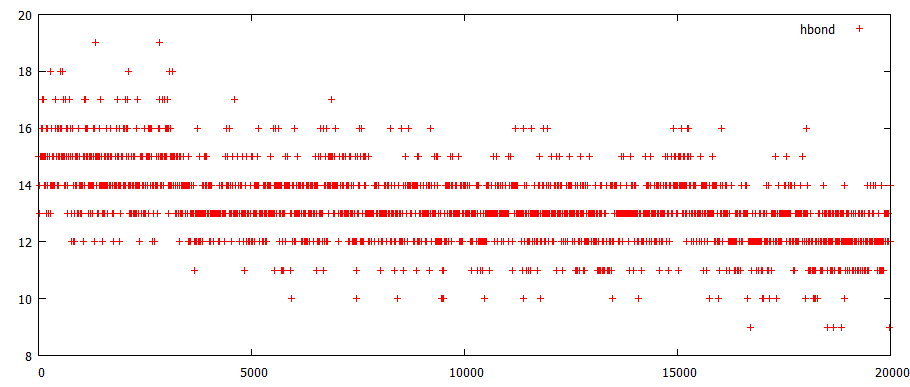
на 0 ns их 15 (хотя для GATCTA последовательности д.б. 14), к 20 ns их остается только 12. Таким образом, разорвалось 3 водородные связи внутри цепей ДНК, что соответствует разрыву комплементарной пары G-С. При этом заметные изменения начинается примерно около 4 ns, что соответствует модели 17-18 - разрушение водородных связей между G-C из-за начала их выхода из одной плоскости, увеличения расстояния между ними.