Анализ результатов плавления ДНК в формамиде
- Общее описание системы
- Детальное исследование результатов моделирования
Силовое поле используемое при построении топологии - amber99sb
Заряд системы - -10 (обусловлен наличием фосфатов в ДНК, которые и вносят отрицательный заряд в систему). Для нейтрализации системы запускаем:
grompp -f em -p dna -c dna_s -o dna_s genion -s dna_s -o dna_si -p dna -np 10
Заряд системы 0. мы это произвели путем внесения 10 положиетльных зарядов в систему.
квадрат 5.01400nm *5.00700nm *5.26800nm- размер ячейки
Минимизация энергии:
integrator = l-bfgs (алгоритм минимизации энергии)
coulombtype = Cut-off (алгоритм рассчета электростатики)
vdwtype = Cut-off (алгоритм рассчета Ван-дер-Ваальсовых взаимодействий)
Описание молекулы растворителя (формамид):
[ moleculetype ] ; molname nrexcl FAM 3 [ atoms ] 1 N 1 FAM N1 1 -0.9108 14.01000 ; amber N type 2 C 1 FAM C2 2 0.6531 12.01000 ; amber C type 3 O 1 FAM O3 3 -0.5717 16.00000 ; amber O type 4 H 1 FAM H11 4 0.4223 1.00800 ; amber H type 5 H 1 FAM H12 5 0.3865 1.00800 ; amber H type 6 H5 1 FAM H2 6 0.0206 1.00800 ; amber H type [ bonds ] 1 2 2 3 1 4 1 5 2 6 [ pairs ] ; ai aj funct c0 c1 c2 c3 3 4 1 3 5 1 6 4 1 6 5 1 [ angles ] ; ai aj ak funct c0 c1 c2 c3 3 2 1 1 3 2 6 1 6 2 1 1 2 1 4 1 2 1 5 1 4 1 5 1 [ dihedrals ] ; ai aj ak al funct definition 3 2 1 4 9 3 2 1 5 9 6 2 1 4 9 6 2 1 5 9 [ dihedrals ] 6 1 2 3 4 2 4 1 5 4
парметр, определяющий фиксацию биополимера: define = -DPOSRES
Утряска растворителя:
количество шагов: 10000
длина шага: 0.001 ps (суммарно - 1.0ps)
Алгоритм расчета электростатики: coulombtype = pme
Алгоритм расчета Ван-дер-Ваальсовых взаимодействий: vdwtype = Cut-off
Алгоритм термостатат: Tcoupl = Berendsen
Алгоритм баростата: Pcoupl = no
Основной расчет МД:
количество процессоров: 16
длина траектории: 20ns
количество шагов: 10000000
длина шага: 0.002 ps
алгоритм интегратора: integrator = md
алгоритм расчета элкутростатики: coulombtype = pme
алгоритм расчета Ван-дер-Ваальсовых взаимодействий: vdwtype = Cut-off
алгоритм термостата: Tcoupl = v-rescale
алгоритм баростата: Pcoupl = Berendsen
Визуальный анализ
dna_pbc_1.pdb- результат визуализации моделирования. Первая нуклеотидная пара нарушается уже на 2ой модели (t=200.00000 фс) На последней моделе (101, t=20000.00000) разрушено всего две нуклеотидные пары, и начинают разрушаться отсальные (выводятся из нормального стэкинг взаимодествия). Видимо, время выбранное для моелирования недостаточно, для осуществления полного плавления олигонуклеотида в формамиде. При этом нарушение второй пары происходит (процесс начинается намного раньше, но полностью пара разрушается лишь к этому моменту времени) на 57 модели (t= 11200.00000).
Средне-квадратичное отклонение
Так как у нас происходит конформационный переход, сначала расчитаем отклонение в ходе всей симуляции относительно стартовой структуры.
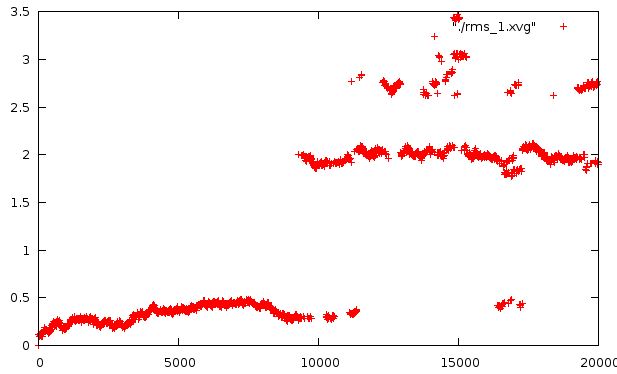
И относительно каждой предидущей структуры на растоянии 400 кадров
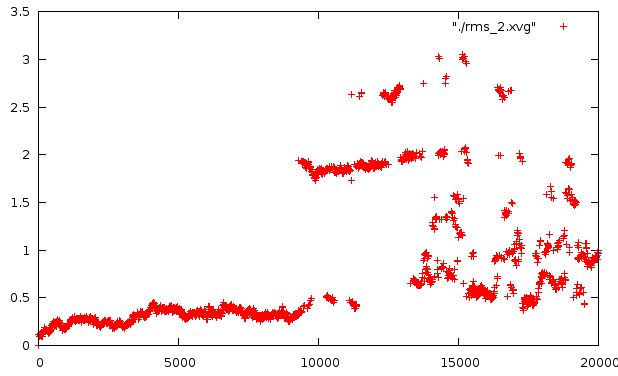
На втором графике видно, что действительно происходит снижение значений, но все-таки они не стабильны, что говорит о неоконченности плавления.
Изменение гидрофобной и гидрофильной поверхности в ходе моелирования
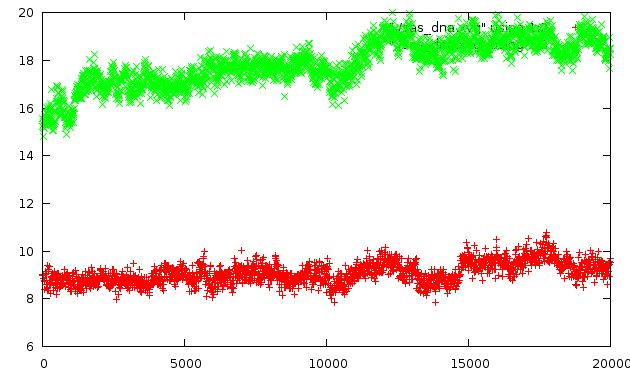
зеленые крестики - гидрофильная поверхность; красные - гидрофобные
из графика видно, что со значениями гидрофобной поверхности значительных изменений не происходит (что и логично), а вот в случае гидрофильной поверхности все гораздо более интересно:
видно, что ее значение изменяется с 15.2297 единиц (начало моделирования) на 18.4922 в конце моделирования (хотя в ходе моделирования наблюдались и большие значения, вплоть до 19.7127 (с чем связано уменьшение этого параметра будет рассмотрено ниже). это происходит в результате вывода гетероциклических оснований (содержащих амино-группы в совем составе) из среды недоступной растворителю (образования водородных связей и стэкинг-взаимодействий между основаниями не позволяет проникать молекулам растворителя) в среду, доступную растворителю.
По этому графику с легкостью можно определить моменты времен распада первой нуклеотидной пары (полностью распадается в районе 1000 фс (наблюдаем резкий скачок)) и распад второй пары (скачок в раоне 11000-12000 фс) (более точно назвать это время не позволяет шкала). Выше описанное падение (с 19.7127 до 18.4922) может быть объяснено следующим фактом: происходит формирование водородных связей между двумя основаниями одной цепи
| модель №90 | модель №100 |
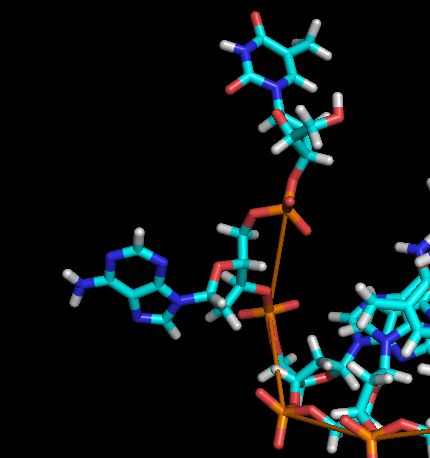
|

|
Водородные связи
На ниже приведенном графике представлена зависимость количество водородных связей в системе от времени моделирования.
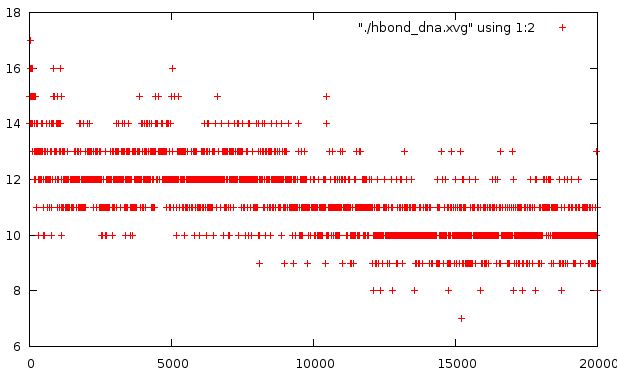
Видно, что в ходе моделирования количество водородных связей снижается (начальное количество 14 - соответствует количеству правильных Н-связей для олига GATCTA). Это также свидетельствует о плавлении ДНК
©Анисенко Андрей