Описание объекта
В данном проекте мы рассматриваем структуру, внутри- и межмолекулярные взаимодействия в мутантной М185С лактальдегид-редуктазе (FucO) принадлежащей E. coli (PDB ID 5BR4, Cahn et al., 2016). Лактальдегид-редуктаза — это фермент, принадлежащий семье оксидоредуктаз (КФ 1.1.1.77). Также фермент известен как пропандиол NAD+ оксидоредуктаза.
Катализируемая реакция:
(R)[or (S)]-propane-1,2-diol + NAD+ → (R)[or (S)]-lactaldehyde + NADH + H+
В процессе реакции диол превращается в лактальдегид, теряя 2 протона и 2 электрона, а NAD+ восстанавливается до NADH. Реакция не является стереоселективной: субстратом может быть как (R), так и (S) изомер пропан-1,2-диола. NAD+ является кофактором. Фермент принимает участие в метаболизме пропановой кислоты[5], глиоксилатном и дикарбоксилатном пути[6].
Описание нативной структуры (PDB ID 1RRM)
Глобулярный белок, присутствующий в цитоплазме бактерий. Молекула является гомодимером из цепей А и В, удерживаемых десятью водородными связями между двумя антипараллельными β-листками, сформированными как цепью А, так и цепью В. Каждая цепь имеет один активный центр с катионом цинка. Рядом расположены сайты связывания субстрата и NAD+. Последний включает в себя DHQS-подобную укладку (DHQS = 3-гидрохинонат-синтаза) — характерную структуру NAD(P)H - связывающих белков, состоящую из чередующихся β-цепей and α-спиралей.
Описание рассматриваемого белка (PDB IB 5BR4) и ее характеристик
Данный мутантный фермент интересен тем, что мутация в его активном центре (M185C) улучшает его каталитическую активность без изменения ее специфичности, структуры белка и других его физико-химических свойств. Обычно мутации, меняющие каталитическую активность ферментов, связаны с изменением активного центра, что может вызывать непредсказуемое изменения сродства к субстрату. Могут затрагиваться даже такие параметры реакции как оптимум pH и температура. Исследования In vitro и in vivo показали[4], что мутация М185С не произвела значительных изменения на характеристики фермента, помимо увеличения скорости катализируемой реакции. Термостабильность не изменилась и бактерии с данной мутацией стали более устойчивы к токсичным альдегидам. Мутация заменяет 185 остаток метионина на цистеин в обеих цепях. Новый остаток расположен дистально к активному центру в DHQS-подобном мотиве NAD+/NADH связывающего сайта. Цистеин является более полярным, чем метионин, и, хотя его боковая цепь почти совпадает с Cα и Cβ атомами метионина, в сайте связывания NAD+/NADH почти не происходит каких-либо больших изменений помимо сдвига аденина NAD-a на 0.3Å по направлению к активному центру. Этот сдвиг приводит к более ощутимому сдвигу N1 атома никотинамида NAD-a на 1.1Å. Поскольку этот атом азота непосредственно участвует в реакции, принимая на себя электроны, предполагается, что его смещение в направлении активного центра играет главную роль в повышении каталитической активности фермента.
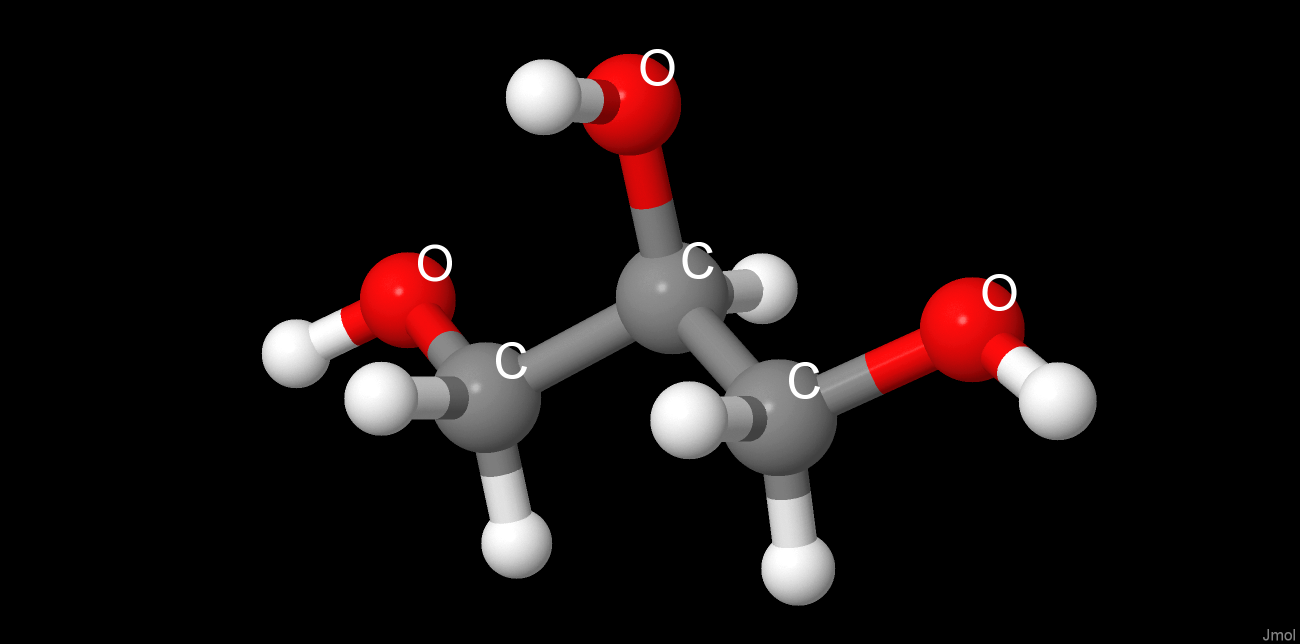
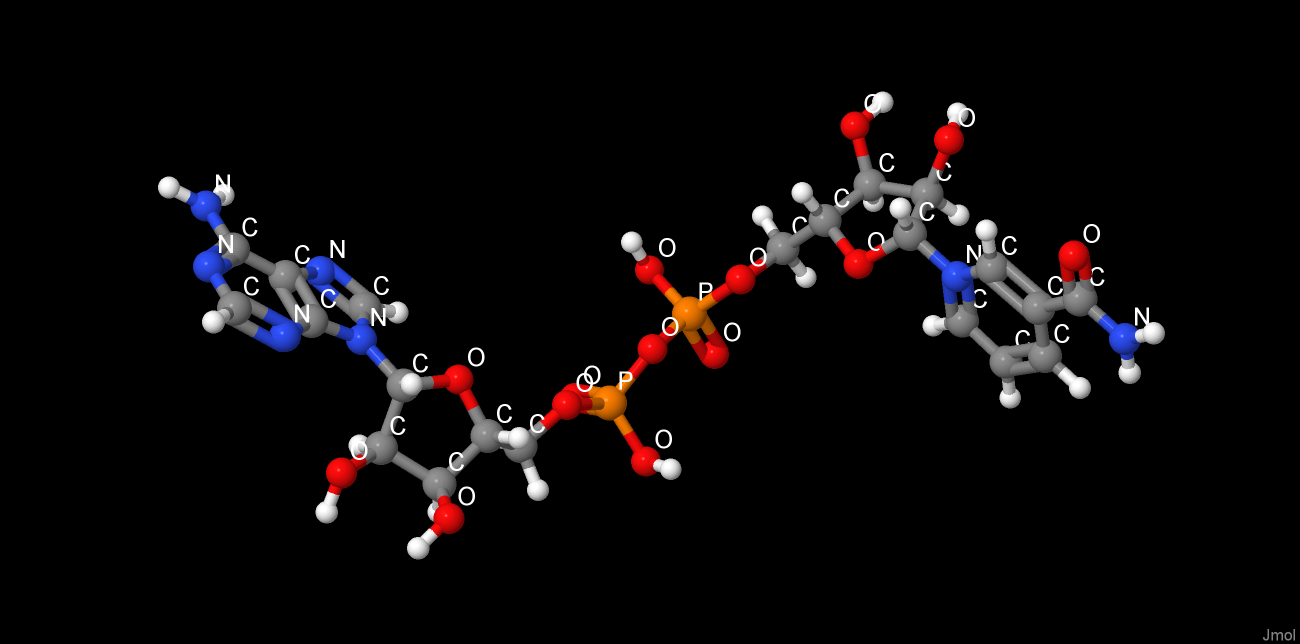
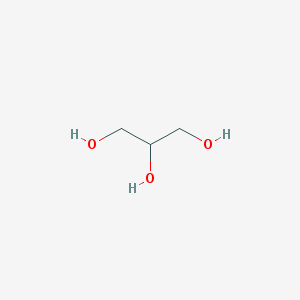
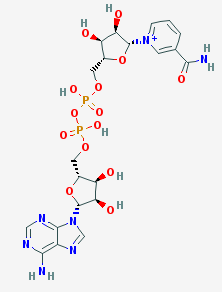
Выбранные методы приготовления кристалла белка[4] значительно изменили четвертичную структуру белка и его лигандный состав. Использование NH4Cl в качестве осадителя вызвало встраивание хлорид-ионов в белок, что не характерно для его нативной структуры. При высушивании белка был использован глицерин. Будучи структурно подобеным пропан-1,2-диолу, глицерин встроился в сайт связывания субстрата. Глицерин изменил четвертичную структуру, встроившись между глобулами и соединив их с помощью образовавшихся водородных связей. В результате цепи соединились со стороны, противоположной наблюдаемой в нативной структуре, разрушив связывающие глобулы β-листки. Таким образом, на один димер в кристалле содержится: две молекулы NAD+, 3 глицерина, из которых 2 занимают активные центры, а 1 является мостиковым, и хлорид-ионы.
Краткая характеристика низкомолекулярных лигандов
|
 |
||||||||||
|
 |
||||||||||
|
 |
||||||||||
|
 |
Описание контактов
Белок-белок
С помощью средств JMol в исходном белке удалось выявить только водородные связи через глицерин. Однако, в водном растворе и, вероятно, in vivo, данный белок имеет другую конформацию в зоне контакта. Мы взяли для определения взаимодействий между цепями другую модель лактальдегид редуктазы в воде (PDB id 1rrm). С помощью команд J(S)Mol contact и hbond calculate было найдено 10 водордных связей, показанных на иллюстрации ниже.
Для примера приведем параметры связей в двух разных листах. Это две идентичные связи, хотя их параметрыи незначительно отличаются.

| Связь | Угол, ° | Длина, Å |
|---|---|---|
| [MSE]6:A.N-[ALA]12:B.O | 165,1 | 2,72 |
| [MSE]6:B.N-[ALA]12:A.O | 164,2 | 2,8 |
Белок-лиганд
Координационные связи с ионом цинка
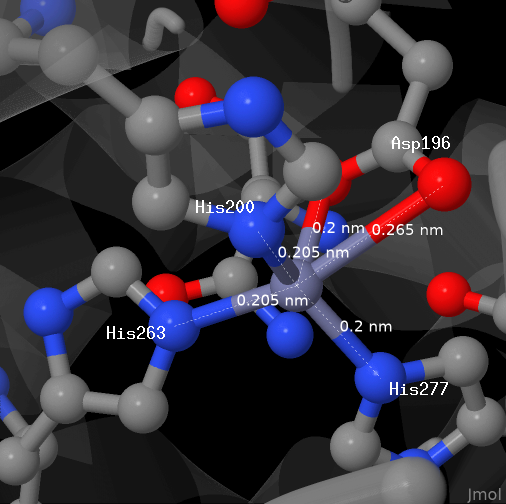
На изображении выше представлен ион цинка в исследуемом белке (цепь А). С ним связано четыре аминокислотных остатка донорно-акцепторной связью. Связь с аспартатом делокализована, поэтому она отображена как две связи, с каждым из двух атомов кислорода. Связи указаны в PDB, отображаются по умолчанию.
Водородные связи НАД
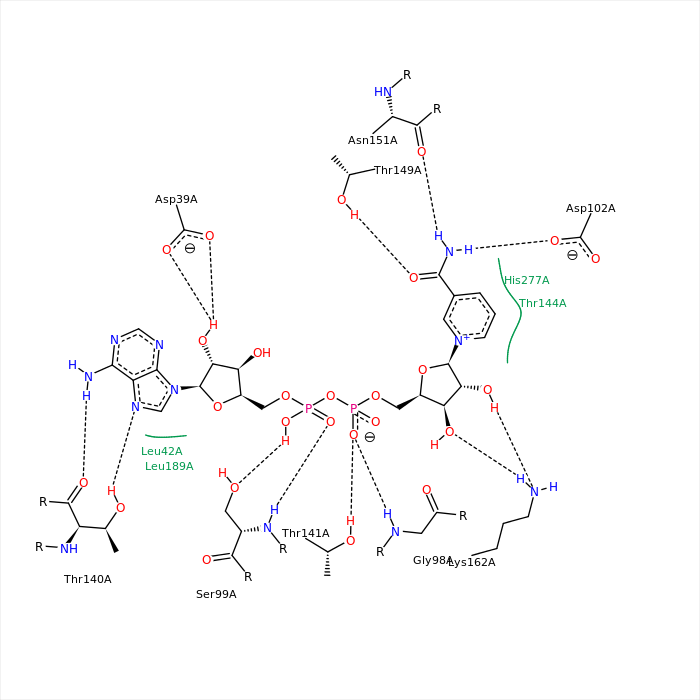
На изображении выше представлен НАД в исследуемом белке (цепь А). Представленная иллюстрация взята с сайта rcsb.org. Водородные связи в J(S)Mol найдены с помощью команды hbonds calculate с предварительно отключенным RasMol. Длины связей колеблются в пределах 2,75-3,24Å. Параметры некоторых связей:
| Связь | Угол, ° | Длина, Å |
|---|---|---|
| [THR]141:A.OG1-[NAD]402:A.O1N | 145,9 | 2,74 |
| [GLY]98:A.N-[NAD]402:A.O1N | 108,8 | 2,85 |

Амино-ароматические взаимодействия
Амино-ароматические взаимодействия очень схожи со стакинг-взаимодействиями и имеют сходную геометрическую конфигурацию. Их суть заключается во взаимодействии δ( + ) боковых цепей остатков с δ(-) π-электронами ароматической системы. Наиболее прочны такие соединения при небольшом пересечении ван-дер-Ваальсовых радиусов взаимодействующих групп, примерно 4,7Å. Наиболее распространен угол с нормалью к плоскости кольца от 30° до 50° у лизина, аргинина, глутамина и аспарагина. Для гистидина еще одна оптимальная область — 80°-90°.
Для примера приведем параметры связи, которая существует в нашем белке: ARG160-PHE272. Для нахождения расстояния и угла воспользовались данными из JMol, по которым был построен треугольник, из которого с помощью простой геометрии вычислили, что длинна связи составляет 3,93Å, а угол равен 23°.

Гидрофобное ядро
Для нахождения гидрофобных ядер воспользовались программой CluD. Выбраны были ядра размером более 5 атомов. Так как исследуемый белок представляет из себя гомодимер, ядра расположены симметрично, по три в каждой цепи.
В качестве MyResidue был выбран остаток лейцина 284:A из гидрофобного ядра №1. Как можно видеть из апплета, атомы, находящиеся на расстоянии 6Å от MyResidue, почти полностью его закрывают. Для определения среднего расстояния между атомами в гидрофобном ядре, не связанными ковалентными связями, мы произвели 20 измерений, по которым была составлена таблица. При расчетах использовались следующие значения ван-дер-Ваальсовых радиусов: углерод - 1,85Å, азот - 1,54Å, кислород - 1,4Å, сера - 1,85Å.[1]
| # | Расстояние между центрами атомов, Å |
Координаты атома #1 | Координаты атома #2 | Расстояние между ван-дер-Ваальсовыми поверхностями атомов, Å |
| 1 | 3,53 | [MET]265:A.CE | [TYR]379:A.CG | -0,17 |
| 2 | 3,88 | [TYR]379:A.CD1 | [LEU]340:A.CD1 | 0,18 |
| 3 | 3,66 | [LEU]340:A.CD2 | [ILE]283:A.CG2 | -0,04 |
| 4 | 4,09 | [VAL]288:A.CG2 | [MET]265:A.SD | 0,39 |
| 5 | 3,85 | [LEU]269:A.CD1 | [LEU]340:A.CD2 | 0,15 |
| 6 | 3,80 | [HIS]287:A.CD2 | [ALA]382:A.CB | 0,10 |
| 7 | 3,38 | [ASN]281:A.O | [LEU]285:A.CB | 0,13 |
| 8 | 3,55 | [ALA]382:A.CB | [LEU]378:A.O | 0,30 |
| 9 | 3,38 | [ILE]283:A.O | [HIS]287:A.CD2 | 0,13 |
| 10 | 3,45 | [ALA]282:A.O | [PRO]286:A.CD | 0,20 |
| 11 | 2,43 | [ASN]281:A.N | [ALA]280:A.CA | -0,96 |
| 12 | 3,85 | [LEU]269:A.CD1 | [LEU]340:A.CD2 | 0,15 |
| 13 | 5,16 | [ALA]382:A.CB | [LEU]284:A.CA | 1,46 |
| 14 | 4,18 | [LEU]284:A.CD1 | [LEU]269:A.CD1 | 0,48 |
| 15 | 4,55 | [LEU]284:A.CG | [LEU]340:A.CD2 | 0,85 |
| 16 | 3,94 | [LEU]284:A.CD1 | [ALA]266:A.CA | 0,24 |
| 17 | 4,25 | [LEU]284:A.CD2 | [ALA]382:A.CB | 0,55 |
| 18 | 4,44 | [LEU]284:A.CD2 | [MET]265:A.SD | 0,74 |
| 19 | 3,82 | [LEU]284:A.CD2 | [MET]265:A.CE | 0,12 |
| 20 | 4,54 | [LEU]284:A.CB | [VAL]288:A.CG2 | 0,84 |
Среднее расстояние между ван-дер-Ваальсовыми поверхностями атомов в гидрофобном ядре составило 0,3Å, чего, конечно же, недостаточно, чтобы разместить между ними атом кислорода радиусом 1,4Å без перекрывания электронных облаков. Среднее межъядерное расстояние составило 3.89Å, допуская перекрывание электронных оболочек, атом кислорода туда войдет.
Вклад авторов
Пушкарев Сергей занимался поиском и описанием гидрофобного ядра, описанием лигандов, написанием и отладкой соответстствующих скриптов, а также сборкой обзорного скрипта и переводом. Бушмакин Илья описывал координационные и водородные связи между белком и лигандами (цинк и НАД), амино-ароматические взаимодействия, писал и отлаживал соответстствующе скрипты, верстал страницу. Пухов Степан изучал литературу, описывал белок, его функции и структуру, поиском альтернативных структур белка и литературы.
Использованная литература
- Дж, Эмсли. Элементы. М.: Мир, 1993.
- Burley, S. K., and G. A. Petsko. "Amino‐aromatic interactions in proteins." FEBS letters 203.2 (1986): 139-143.
- Dougherty, Dennis A. "Cation-π interactions in chemistry and biology: a new view of benzene, Phe, Tyr, and Trp." Science 271.5246 (1996): 163-168.
- Cahn, J.K.B. et al. “Mutations in Adenine-Binding Pockets Enhance Catalytic Properties of NAD(P)H-Dependent Enzymes.” Protein Engineering, Design and Selection 29.1 (2016): 31–38. PMC. Web. 7 Mar. 2018.
- "KEGG PATHWAY: map00640",KEGG: Kyoto Encyclopedia of Genes and Genomes, http://www.genome.jp/kegg.
- "KEGG PATHWAY: map00630",KEGG: Kyoto Encyclopedia of Genes and Genomes, http://www.genome.jp/kegg.