- Краткое описание структуры в файле 1PP7.pdb С
сайта PDB я скачала файл
1PP7.PDB,в котором приведены координаты атомов следующих молекул:
- 39 kDa initiator binding protein (из простейшего Trichomonas vaginalis) - 1 молекула
- Ferredoxin инициаторный элемент(Inr, ДНК, искусственно синтезирована) - 1 молекула (2 цепи)
- ионы цинка (Zn+2) - 7 ионов
- Вода - 65 молекул
F 5' [15] CAAGTGAAGTAAC [3] 3' | |||||| | | E 3' [25] GGTTACTTCACTT [37] 5'
Картинка ДНК-белкового комплекса:
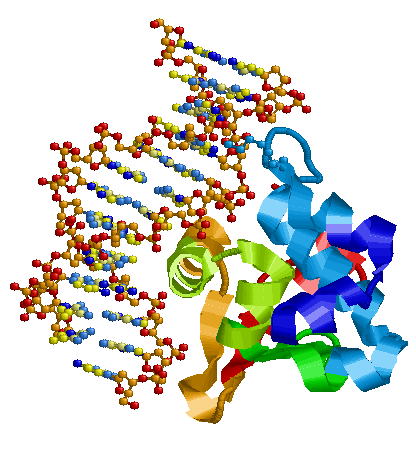
- Функции белка, структура которого представлена в файле 1PP7.pdb
В соответствующем документе UniProt, можно найти описание функций
(и особенностей) белка:
Код доступа ("Accession number") Q95VR4 Идентификатор записи в БД Q95VR4_TRIVA Название (краткое описание) белка 39 kDa initiator binding protein
EC=6.1.1.-;Длина: 341 AA Организм: Trichomonas vaginalis Идентификаторы записей PDB 1PP7; 1PP8; 1Q87; 1Q88; 1Q89 Примечания: Статус белка в базе: Unreviewed - Исследование структуры ДНК
С помощью программ find_pair и analyze (выходной файл)
определен тип формы ДНК: правозакрученная B-ДНК. Так же с помощью Excel были определены средние значения торсионных углов
для внутренних нуклеотидов (для всех, кроме краевых) и номер самого "деформированного" нуклеотида (с самой большой суммой модулей торсионных углов)
- С34 на цепи Е (файл (torsion_DNA.xls) с рассчетами).
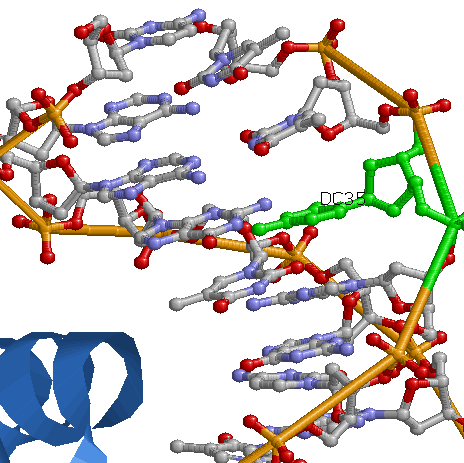
Самый "кривой" нуклеотид с виду не радикально отличается от остальных, не близок к белку, чем можно было бы объяснить искривление. - Исследование природы ДНК-белковых контактов
Скрипт my_dna.def
Таблица. Контакты разного типа в комплексе 1PP7.pdb
Из таблицы видно, что больше всего - неполярнх контактов белка с сахарофосфатным остовом ДНК. Это вполне закономерный результаты: остовные атомы находятся на поверхности молекулы ДНК, а большее количество неполярных контактов объясняется тем, что в белке вообще больше неполярных атомов, и тем, что расстояние для неполярного контакта может быть больше, что приводит к большей вероятности такого контакта.Контакты атомов белка с Полярные Неполярные Всего остатками 2'-дезоксирибозы 2 22 24 остатками фосфорной кислоты 4 11 15 остатками азотистых оснований со стороны большой бороздки 1 - 1 остатками азотистых оснований со стороны малой бороздки - 5 5
Остатки азотистых оснований находятся внутри молекулы, что делает их труднодоступными для контактов. Немного больше контактов у атомов, обращенных в сторону маленькой бороздки - это связано либо с тем, что маленькая бороздка мельче, а значит, атомы в ней расположены ближе к поверхности, либо просто со статистическими погрешностями - случайным преобладанием таких контактов (участок ДНК для анализа очень небольшой).
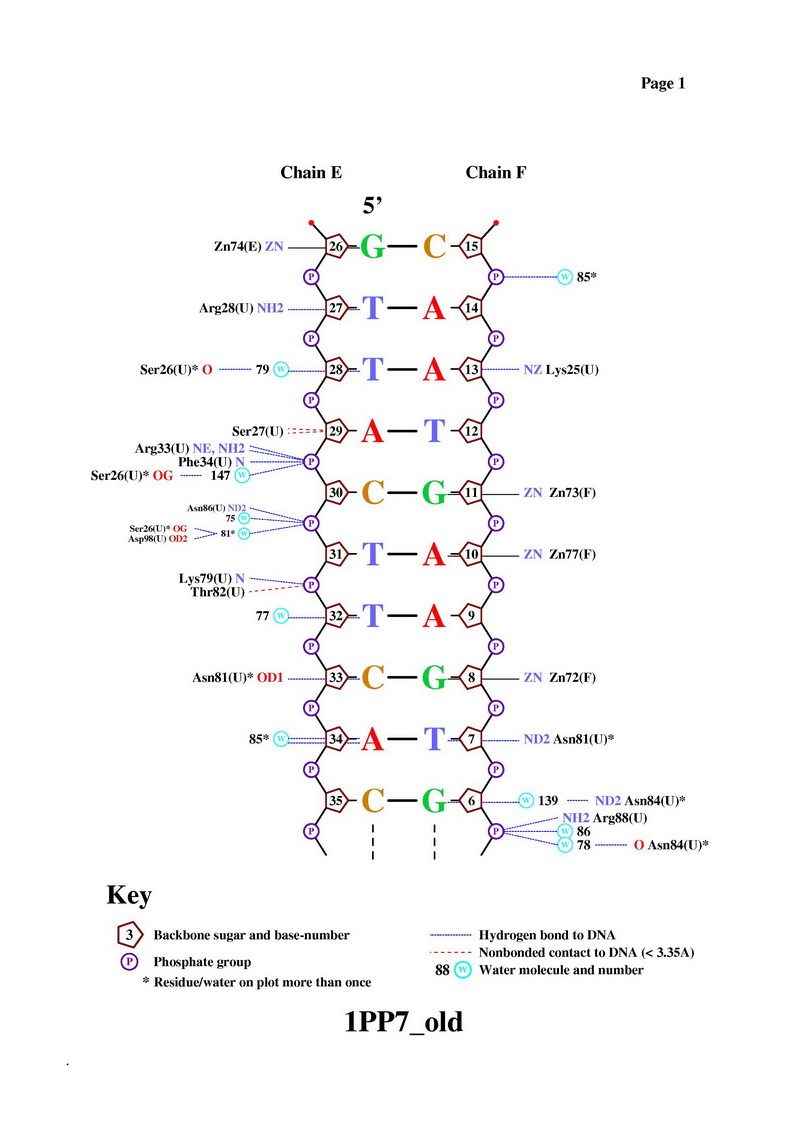
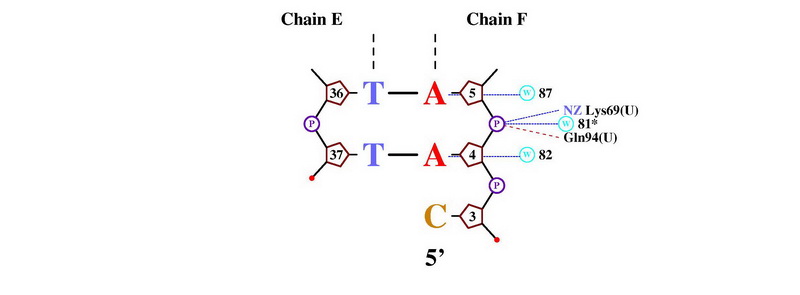
- Получение популярной схемы ДНК-белковых контактов с помощью nucplot
С помощью команды nucplot 1PP7.PDB был получен файл в формате .ps, который был переведен в картинку JPEG:
- Возможный(е) распознающий(е) контакт(ы)
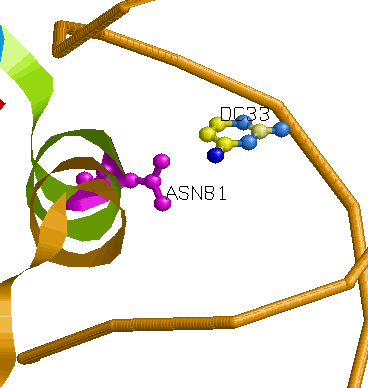
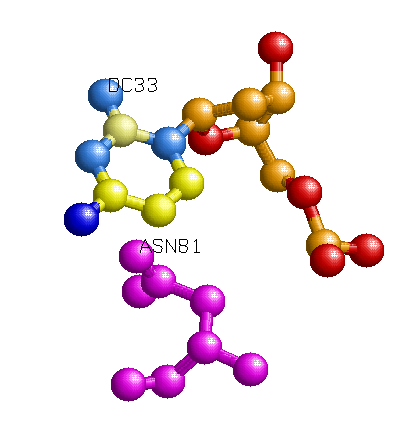
Одним из наиболее вероятных распознающих контактов мне кажется контакт Asn81 и С33 (на рисунке выше). Во-первых, на популярной схеме контактов помечено, что между аспарагином и цистеином более одного контакта, во-вторых, это один из небольшого числа прямых контактов между аминокислотным остатком и остатком азотистого основания (что придает большую специфичность контакту, чем контакт с остовными атомами). Также на изображении RasMol видно, что спираль, из которой "торчит" Asn81 практически лежит в бороздке, что создает специфичность контакта на уровне формы третичной структуры молекул. Изображение контакта:
- Характеристика ДНК-связывающего домена Q95VR4_TRIVA
С помощью инструментов Pfam определила доменную структуру белка из исследуемого комплекса:
Transcription-initiator DNA-binding domain IBD

(Не найден в InterPro)
© Smirnova Victoriya, 2009