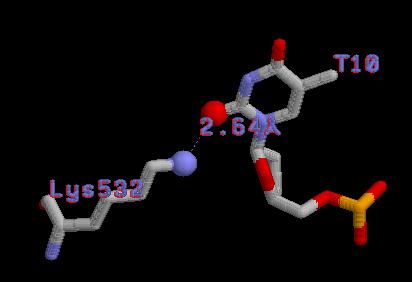
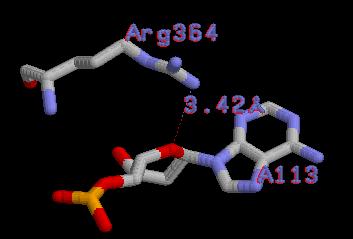
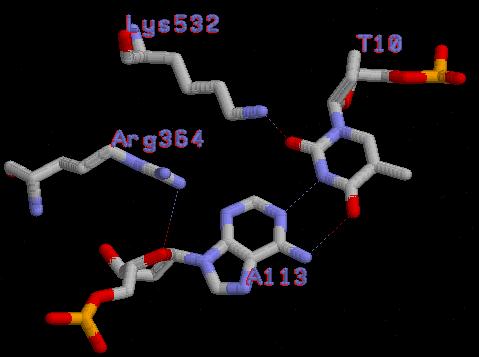
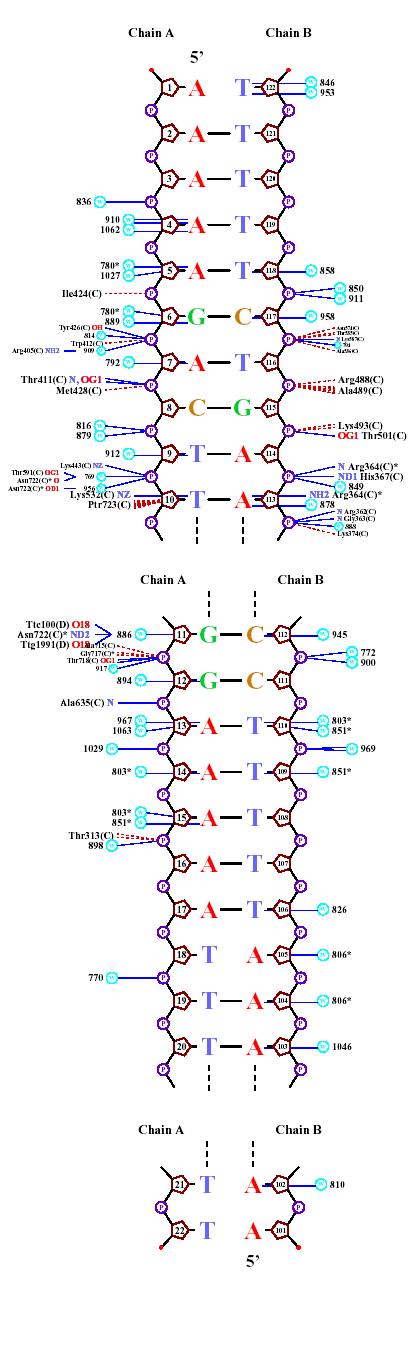
| Контакты белка с... | Полярные | Гидрофобные | Всего |
| ...остатками 2`-дезоксирибозы | 6 | 11 | 17 |
| ...остатками фосфорной кислоты | нет | нет | нет |
| ...остатками азотистых оснований со стороны большой бороздки | 40 | 1 | 41 |
| ...остатками азотистых оснований со стороны малой бороздки | 2 | нет | 2 |