Автор: Литвинова Е.Ю.
Руководители: Спирин С.А., Алексеевский А.В.
В работе произведен поиск ортологов гомеобелков семейства "paired" у Drosophila melanogaster и Anopheles gambiae.
Гомеодомен - это консервативный ДНК-связывающий домен, входящий в состав многих эукариотических факторов транскрипции [1]. Гомеобелки - это белки содержащие гомеодомены. Они участвуют в регуляции ключевых процессов развития и гомеостаза. Гомеодомены состоят примерно из 60 аминокислотных остатков, в белке они обычно расположены вблизи C-конца.
Существуют различные классификации гомеодоменов [2]. При классификации учитывается степень сходства последовательностей гомеодоменов, присутствие других белковых доменов в гомеобелке и наличие тех или иных характерных последовательностей аминокислот вблизи гомеодомена. Данная работа проводилась на гомеодоменах семейства "paired".
Белки, принадлежащие к классу paired влияют на первичные функции развития. Они определяются присутствием гомеодомена, сходного с кодируемым Drosophila prd геном, и характеризуются неизменными аминокислотными остатками, отличающими их от других классов гомеодоменов.
Гомеодомен, кодируемый геном Drosophila prd является основополагающим в классе paired, также парный белок (кодируемый Drosophila prd геном) был первым, в котором был обнаружен другой тип ДНК-связывающей структуры - парный домен.
В гомеодомене класса paired должно присутствовать по крайней мере 5 из 6-ти диагностических аминокислотных остатков: P26, D27, E32, R44, Q46, A54.
Традиционно гомеобелки класса paired делят на три подкласса, различающиеся по 50-й позиции гомеодомена, которая играет ключевую роль в определении ДНК-связывающей специфичности.
PAX или prd-type имеют гомеодомен с серином в 50-й позиции и второй ДНК-связывающий - парный домен.
K50 prd-like характеризуются серином в 50-й позиции. У них отсутствует парный домен.
Q50 prd-like содержат глутамин в 50-й позиции и у них также отсутствует парный домен.
Но исследования последовательностей гомеодоменов, проведенные Brigitte Galliot и др. [3], показали, что филогенетический анализ не поддерживает данную классификацию, поэтому она была несколько пересмотрена. Они делят класс paired на 2 подкласса: Prd-like тип, включающий K50 и Q50, и Pax-type, который содержит S50 и парный домен .
Плодовая мушка Drosophila melanogaster и малярийный комар Anopheles gambiae были выбраны в качестве объектов исследования по причине того, что их геномы полностью секвенированы[4,5]; кроме того большое количество белков D.melanogaster аннотированы в банке Swiss-Prot.
Цель работы - найти парные гомеодомены в геноме комара, проследить эволюцию гомеодоменов гомеобелков семейства paired и сравнить доменный состав полных белков.
В работе использовались данные баз SwissProt, TrEMBL [6], Anobase, Pfam. Также были использованы программы пакета EMBOSS [7]: ehmmbuild, ehmmsearch, emma, eneighbor, eprotdist и сервисы SRS и DotHelix.
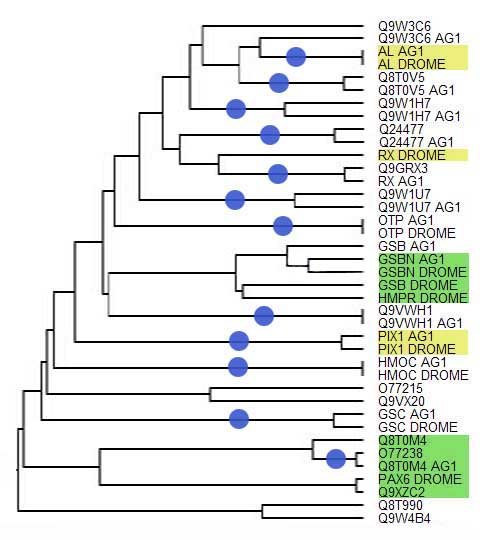
Различие в количестве гомеодоменов у D.melanogaster и у A.gambiae, вероятно,
свидетельствует о произошедших эволюционных изменениях, таких как дупликация или потеря генов.
Анализируя дерево, можно выделить два эволюционных пути расхождения гомеодоменов
D.melanogaster и A.gambiae.
Первый путь - эволюция гомеодоменов в двух разных организмах от единого
предшественника с сохранением общности функции, без дупликаций генов.
На филогенетическом дереве о таком пути свидетельствует наличие отдельных
ветвей, заканчивающихся парой листьев, на одном из которых гомеодомен
D.melanogaster, а на другом A.gambiae . При этом, чем более удален узел
с этими листьями от других узлов, тем более вероятна ортологичность доменов.
Всего было обнаружено 12 пар ортологов.
Отметим, что эти пары ортологов эволюционировали с разной скоростью.
Было обнаружено 4 пары, скорость эволюции которых нулевая, то есть
гомеодомены D.melanogaster и A.gambiae абсолютно идентичны и не
произошло ни одной мутации относительно общего предшественника.
У оставшихся 8 пар скорость эволюции отличается от нуля, то есть
гомеодомены несколько отличаются друг от друга, но меньше, чем от остальных.
Второй путь - эволюция гомеодоменов с возможными дупликациями,
потерями генов. В этом случае на дереве гомеодомены
D.melanogaster и A.gambiae образуют группы, в которых нельзя
выделить явных ортологов. Такие группы содержат неравное число
гомеодоменов исследуемых организмов, например, 3 гомеодомена
D.melanogaster и 2 гомеодомена A.gambiae.
К тому же нельзя исключить неправильное предсказание кодирующих участков,
ошибки секвенирования и т. д. (которые вероятнее встретить в недавно
секвенированном геноме A.gambiae, чем в геноме хорошо изученного
объекта - D.melanogaster), однако даже такими ошибками нельзя полностью
объяснить наблюдаемую картину.
Парное выравнивание полных гомеобелков показало, что участок гомологии обычно выходит за границы гомеодомена. Домены OAR и PAIRED BOX, которые есть в белках D.melanogaster, присутствуют и в белках A.gambiae, гомеодомены которых ортологичны соответсвующим D.melanogaster. Кроме этих аннотированных доменов в большинстве исследованных белков присутствуют и другие неаннотированные участки гомологии.
Дерево гомеодоменов
Желтым выделены белки, у которых есть домен OAR;
зеленым выделены белки, у которых есть парный домен;
синие кружки обозначают пары предполагаемых ортологов.