1. Исследование дисульфидных связей в белке 1ST9_BACSU
Дисульфидная связь (мостик) - это ковалентная связь между двумя атомами серы аминокислоты цистеина.
Она образуется в результате посттрансляционной модификации белка - окисления цистеина и его димеризации. Образуется некодируемая аминокислота цистин. Посттрансляционная модификация цистеинсодержащих пептидов регулирует их биологическую активность. Цистин содержится в основном в белках, секретирующихся из клетки.
Дисульфидные связи поддерживают вторичную и третичную структуры белка. Например, они обуславливают физические свойства кератина.
Для сравнения, инсулин содержит две полипептидные цепи, соединенные дисульфидными мостиками.
(Не привожу картинки сульфидных связях в инсулине, достаточно и демонстрации связей в моём белке)
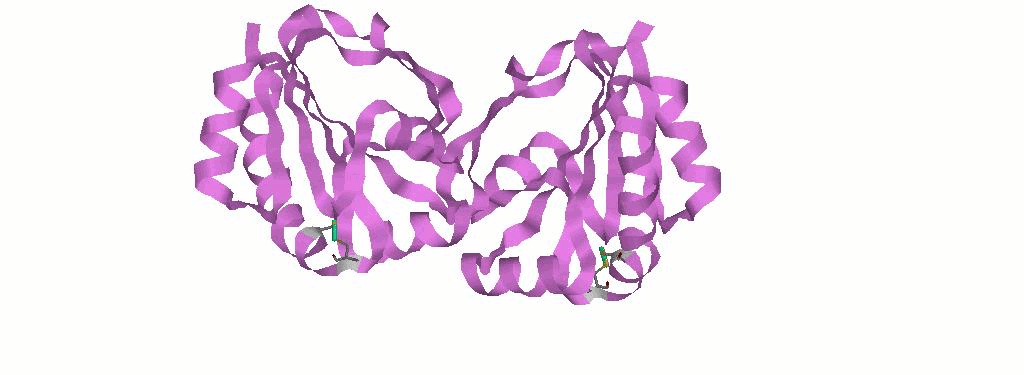
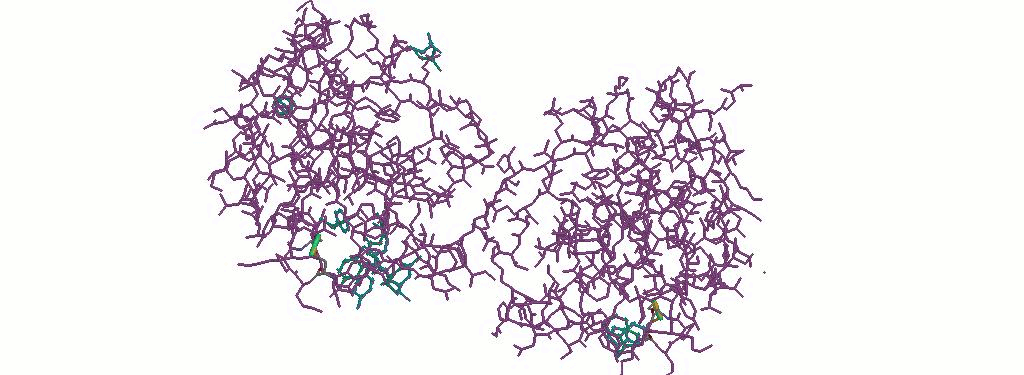
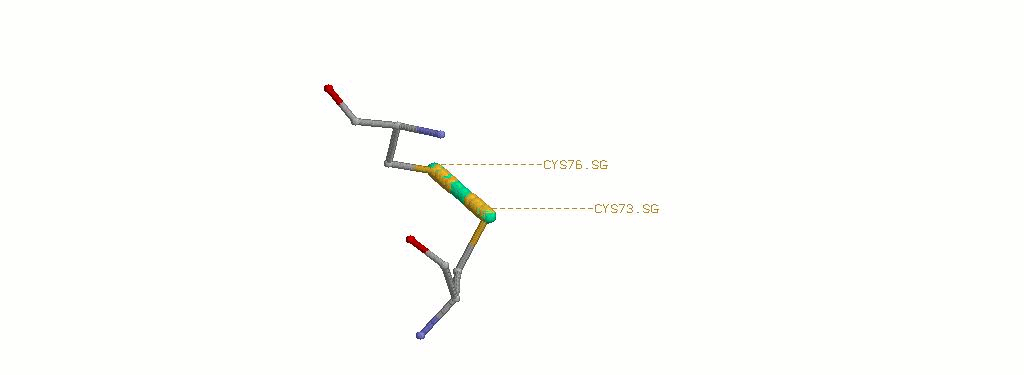
В данной мне структуре белка имеется целых 2 S-S связи:
CYS73A.SG-CYS76A.SG
CYS73B.SG-CYS76B.SG
1.1 Дисульфидные свзяи в белке - вид белка в ленточной модели
1.2 Дисульфидные связи в белке - вид белка в проволочной модели
1.3 Дисульфидная связь в увеличении
2. Исследование контактов с лигадами
В исследуемом белке имеется 4 лиганда этандиола, три из которых расположены в цепи A, один - в B.
На расстоянии меньшем 3.2, удоволетворяющем условию водородной связи, около лигандов расположены части радикала положительно заряженного остатка лизина и молекулы воды. Я предполагаю, что молекулы EDO501A, EDO502A, EDO503B удерживаются водородными связями концевыми атомами азотов остатков лизинов.
Но EDO504A, кажется, "летает в воздухе". На расстоянии 5 ангстрем от него расположены только молекулы воды, один остовный азот, несколько также остовных кислородов, довольно много углеродов и кислород радикала серина 39 на расстоянии 4.18 ангстрем - самый ближний атом к лиганду. Предположить гидрофобное взаимодействие невозможно, хотя бы потому, что этандиол - полярная молекула:
2.1 Этандиол - лиганд

До водородной связи или ван-дер-ваальсова взаимодействия расстояние явно не дотягивает ещё 1 ангстрем. Можно предположить, что в общем случае этандиол удерживается силами электростатического притяжения.
Не буду о грустном и потому рассмотрю предположительно водородное взаимодействие лизина и EDO502A.
2.2 Окружение EDO502, в т.ч. атомы воды

2.3 Окружение EDO502
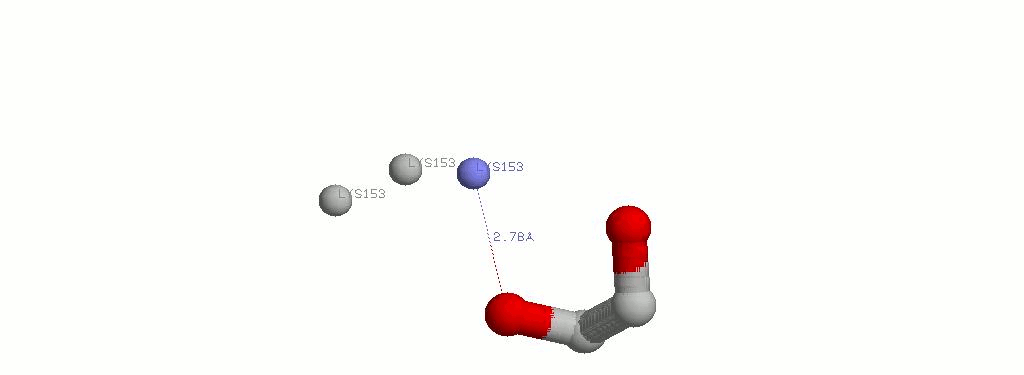
2.4 EDO502A и LYS153A
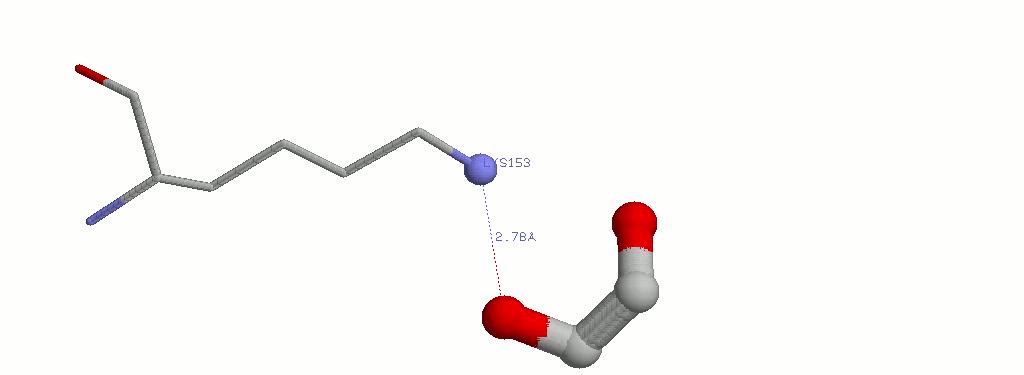
Таблица: Контакты этандиола с белком 1ST9_BACSU
| Имя атома контакта | Имя контактирующего атома белка | Расстояние в Å | Предположительная природа контакта |
| EDO502A.01 | LYS153A.NZ | 2.78 | Водородная связь |
| EDO502A.02 | LYS153A.NZ | 2.96 | Водородная связь |
| EDO501A.01 | LYS60A.NZ | 2.75 | Водородная связь |
| EDO503A.01 | LYS60B.NZ | 2.68 | Водородная связь |
| EDO503A.02 | LYS60B.NZ | 3.23 | Водородная связь |
| EDO504B.01 | SER39A.OG | 4.18 | Электростатическое притяжение ? |
| EDO504B.02 | SER39A.OG | 5.34 | Электростатическое притяжение ? |
3. Описание гидрофобных контактов.
Начальная информация.
Для нашего случая гидрофобность - это свойство аминокислотных остатков неполярных аминоксилот (а.о.) избежать контакта с водой. Из-за гидрофобности а.о. неполярных аминокислот вворачиваются внутрь белка и образуют гидрофобные участки белка, связанные гидрофобными взаимодействиями.
Взаимодействия гидрофобных участков обуславливают свойства шаперонов: шапероновый комплекс имеет на внутренней поверхности участки, обогащенные гидрофобными радикалами аминокислот. К ним присоединяются частично денатурировавшие (развернувшиеся) белки цитозоля с открытыми гидрофобными последовательностями, далее шапероны осуществляют правильное сворачивание белков.
Неполярные гидрофобные аминокислоты:
ALA, ISO, LEU, MET, PHE, PRO, TRY, VAL
Для удоволетворения научного интересна я рассмотрела гидрофобные контакты и фенилаланина, и лейцина в данном мне белке (который, вообще-то, является обычным ферментом).
3.1 Описание гидрофобных контактов фенилаланина
3.1.1 Все фенилаланины с атомами из радиуса 3.5 ангстрем.
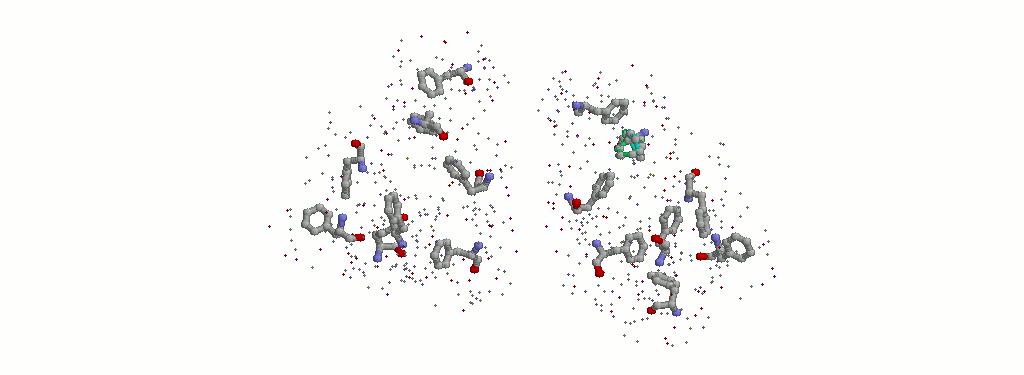
Визуально выбрала фенилаланин с самым <густым> окружением. Им оказался PHE112B.
Его окружают атомы аминокислот:
MET113, TRP69, PHE80, ALA108, VAL109, THR71, ACN111, SER115, LYS77, TYR116, HIS110
Из них гидрофобными являются:
MET113, TRP69, PHE80, ALA108, VAL109.
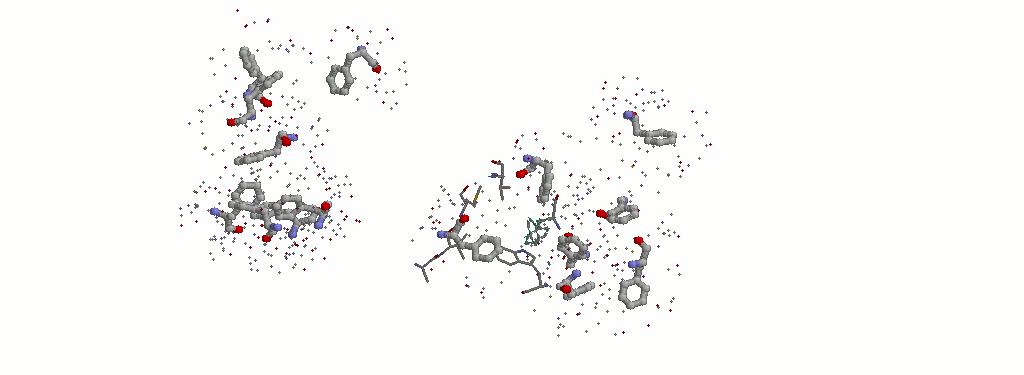
3.1.2 Все фенилаланины с атомами гидрофобных аминокислот из радиуса 3.5 ангстрем. Выделены аминокислоты, окружающие PHE112B.
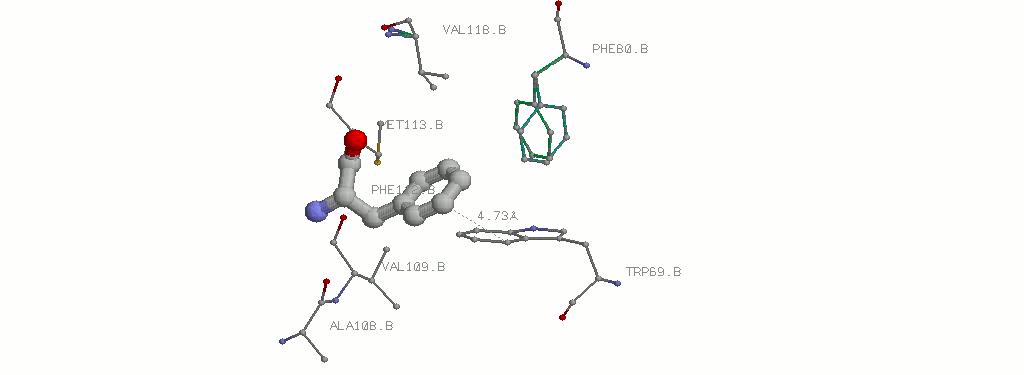
3.1.3 PHE112B и окружающие его гидрофобные аминокислоты. Показано наименьшее расстояние.
3.2 Описание гидрофобных контатов лейцина
3.2.1 Все лейцины с атомами из радиуса 3.5 ангстрем.
Визуально выбрала лейцин с самым <густым> окружением. Им оказался LEU131A.
Его окружают атомы аминокислот:
ALA133, PHE68, VAL130, TYR134, VAL136, VAL102, ASP125, ARG128, ASP135, ASP132
Из них гидрофобными являются:
ALA133, PHE68, VAL130, TYR134, VAL136, VAL102.
3.2.2 Все лейцины с атомами гидрофобных аминокислот из радиуса 3.5 ангстрем. Выделены аминосилоты, окружающие LEU131A.
3.2.3 LEU131A и окружающие его гидрофобные аминокислоты. Показано наименьшее расстояние.
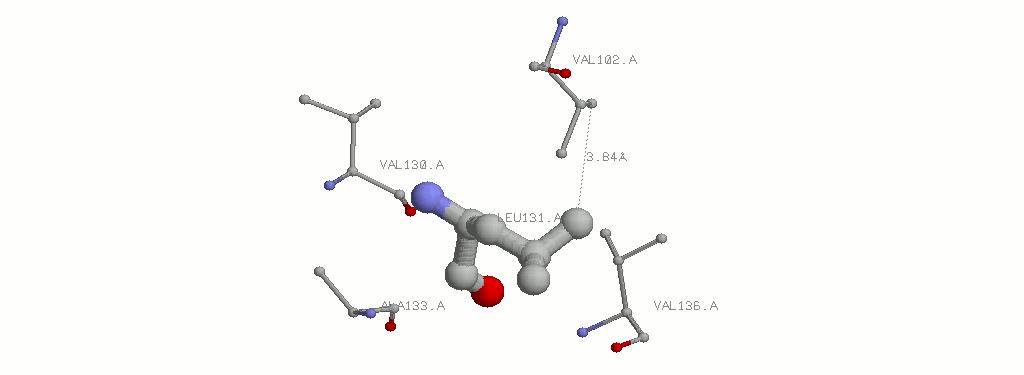
Как видно из картинок, ничего принципиально различного в гидрофобных контактах лайцина и фенилаланина нет, научный интерес не удоволетворен.
Продолжим исследование связей.
4. Водородные связи радикалов.
С теоретической основой здесь, вроде бы, всё ясно.
4.1 Водородные связи радикалов типа O-H... :O.
Для их поиска, выделила весь белок в проволочной модели. Выбрала все атомы кислорода, не входящие в его остов. Раскрасила всё по аминокислотам.
4.1.1 Весь белок в проволочной модели, шарики - атомы кислорода.
Теперь удобно, потыкав мышкой, найти хотя бы одну водородную связь между атомами кислорода радикалов разных аминокислот (т.е. расстояние между аминокислотами не более 3.5 ангстрем). Не будем заморачиваться с тем, кто является донором, а кто - акцептором.
4.1.2 Выбранная водородная связь в приближении. Указано расстояние между атомами кислорода.
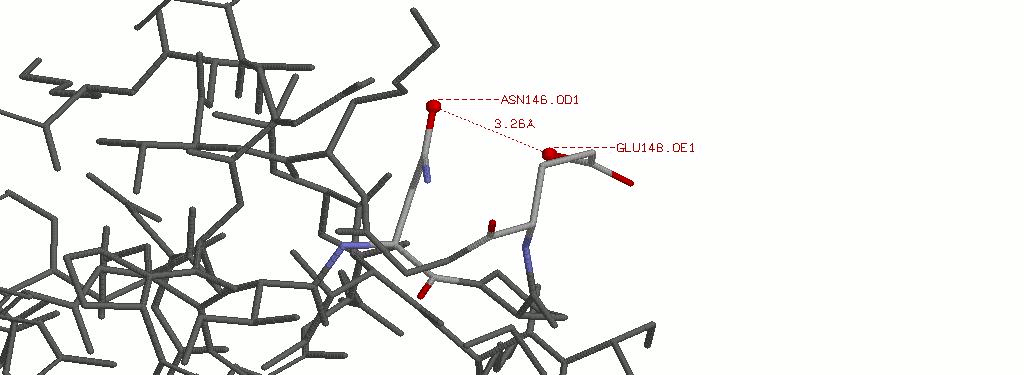
4.2 Водородные связи радикалов типа N-H... :N
Провела все те же действия, что и с кислородом, только с азотом.4.2.1 Весь белок в проволочной модели, шарики - атомы азота
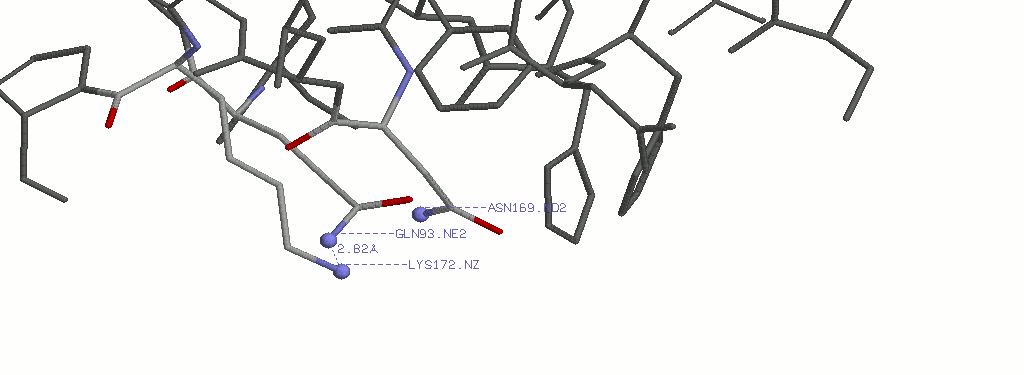
4.2.2 Выбранная водородная связь в приближении. Указано расстояние между атомами азота.
Главная страница
© Галицына Александра, 2011