В данной работе проводится анализ нековалентных взаимодействий в белковой структуре
PDB 2X42 —
бета-глюкозидазе 3B из Thermotoga neapolitana (мутант D242A).
Белок состоит из 715 видимых в кристалле аминокислотных остатков цепи A (полная последовательность — 721 остаток) и относится к семейству гликозидгидролаз GH3 с трёхдоменной архитектурой: N-концевой (β/α)₈-баррель, α/β-сэндвич и C-концевой домен фибронектина III.
Метод получения — рентгеноструктурный анализ (разрешение 2.10 Å, pH 7.4).
Все изображения получены в программе PyMOL с использованием команды ray.
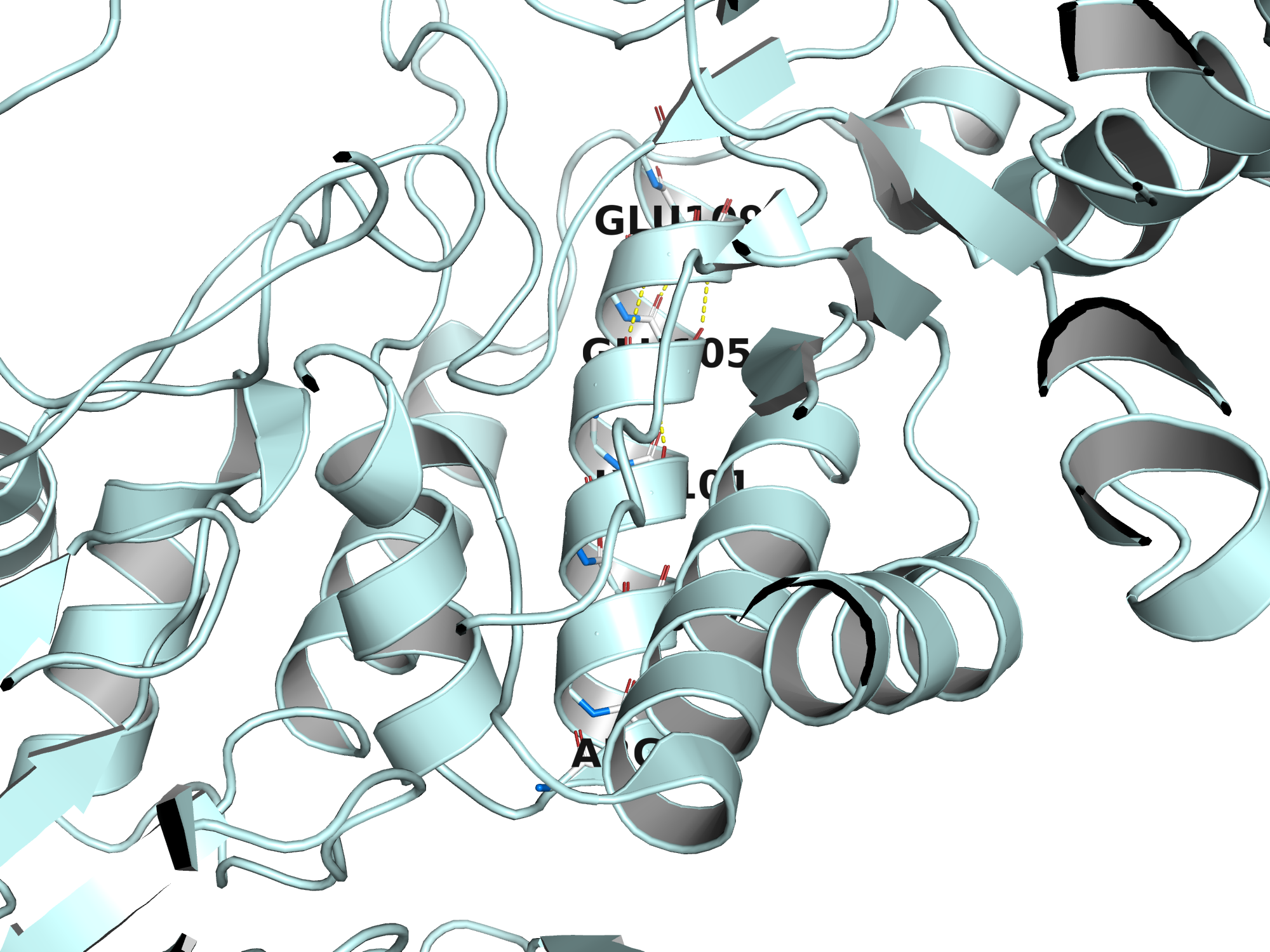
Водородные связи остова (N–H···O=C пептидной группы) формируют вторичные структурные элементы белка. В правой α-спирали каждый донор N–H остатка i образует водородную связь с карбонильным кислородом остатка i−4.
Для иллюстрации выбран участок α-спирали (остатки 92–110 цепи A, HELIX 5 в PDB), расположенной в N-концевом TIM-барреле. Атомы остова показаны палочками: синий — азот (N), красный — кислород (O), белый — углерод (C). Жёлтые штриховые линии — водородные связи N···O.
| Донор (N) | Акцептор (O) | d, Å | Δi |
|---|---|---|---|
| N(104) | O(100) | 2.70 | i−4 |
| N(105) | O(101) | 2.82 | i−4 |
| N(107) | O(103) | 2.83 | i−4 |
| N(108) | O(104) | 2.87 | i−4 |
| N(97) | O(93) | 2.88 | i−4 |
| N(109) | O(105) | 2.93 | i−4 |
Все значения расстояний (2.7–3.0 Å) и паттерн i→i−4 однозначно подтверждают α-спиральную конформацию данного участка.
Водородные связи между боковыми цепями играют ключевую роль в связывании субстратов. Для иллюстрации выбрана область активного центра: молекула α-D-глюкопиранозы (GLC) связывается в подсайте −1 через водородные связи с боковыми цепями нескольких остатков.
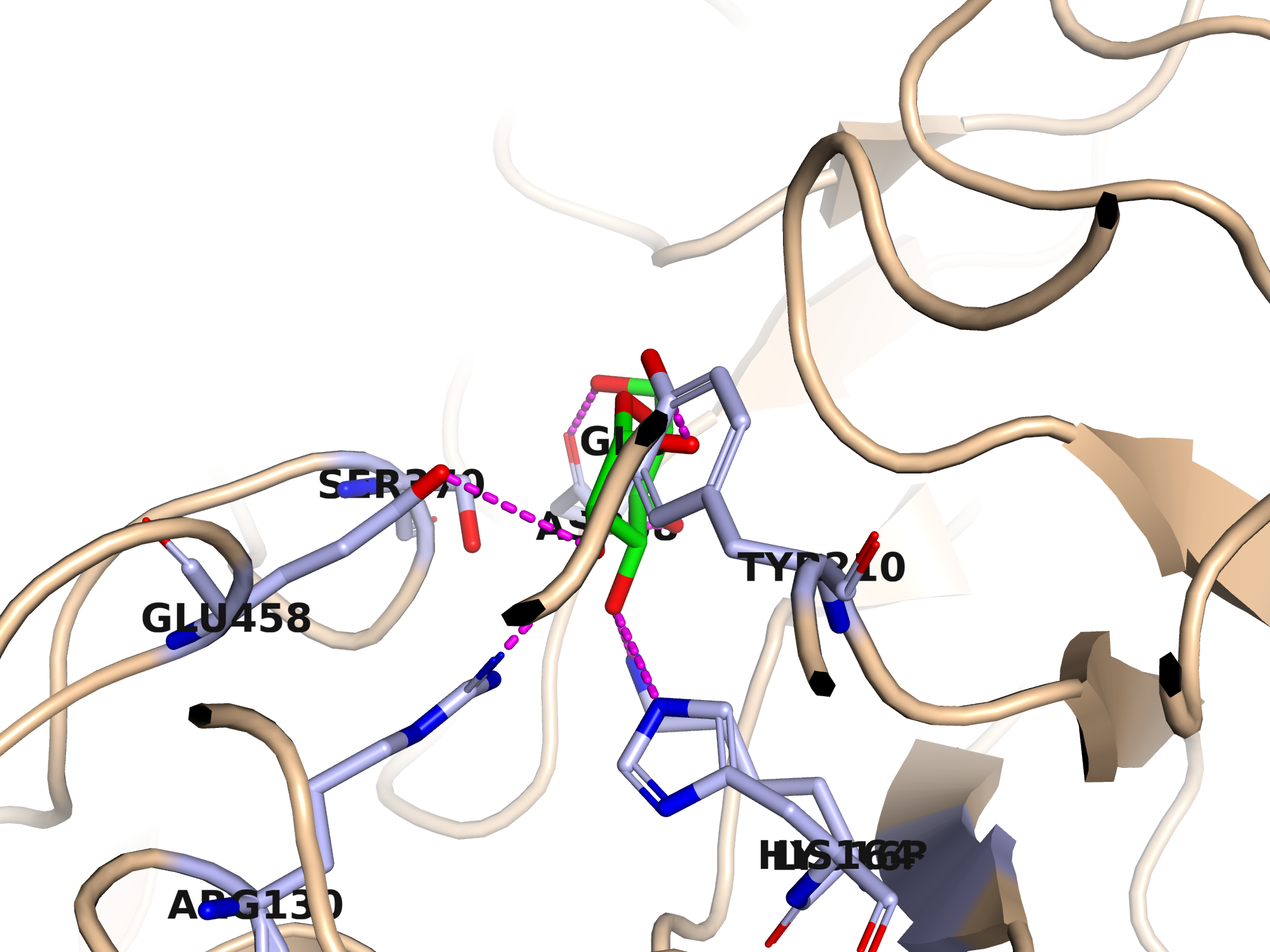
| Остаток | Атом белка | Атом GLC | d, Å |
|---|---|---|---|
| Asp58 | OD1 | O6 | 2.53 |
| Asp58 | OD2 | O4 | 2.58 |
| Lys163 | NZ | O3 | 2.79 |
| Arg130 | NH1 | O2 | 2.81 |
| His164 | NE2 | O3 | 2.81 |
| Tyr210 | OH | O1 | 2.88 |
| Glu458 | OE2 | O2 | 3.34 |
Наиболее короткие водородные связи образует Asp58 (2.53–2.58 Å) — ключевой остаток подсайта −1. Glu458 — каталитический кислотно-основной остаток GH3; мутант D242A в данной структуре лишён нуклеофила.
Солевой мостик — электростатическое взаимодействие между отрицательно заряженными (Asp, Glu) и положительно заряженными (Arg, Lys) боковыми цепями (d ≤ 4.0 Å). В структуре 2X42 обнаружено 64 уникальных солевых мостика — значительное число, характерное для термостабильных белков T. neapolitana (оптимум роста ≈ 90 °C).


| Остаток (−) | Остаток (+) | dmin, Å |
|---|---|---|
| Glu395 | Lys444 | 2.49 |
| Glu232 | Arg187 | 2.56 |
| Asp58 | Arg64 | 2.58 |
| Glu387 | Arg527 | 2.60 |
| Glu686 | Arg718 | 2.64 |
| Glu666 | Arg660 | 2.65 |
| Glu475 | Lys427 | 2.66 |
| Glu334 | Lys632 | 2.71 |
Дисульфидная связь образуется между атомами Sγ двух цистеинов (d(S–S) ≤ 2.5 Å). В структуре 2X42 четыре остатка цистеина: Cys128, Cys161, Cys218 и Cys296. Записи SSBOND в PDB-файле отсутствуют.

| Пара цистеинов | d(Sγ···Sγ), Å | Вывод |
|---|---|---|
| Cys128 — Cys161 | 17.55 | нет связи |
| Cys128 — Cys218 | 14.02 | нет связи |
| Cys128 — Cys296 | 28.67 | нет связи |
| Cys161 — Cys218 | 13.48 | нет связи |
| Cys161 — Cys296 | 15.54 | нет связи |
| Cys218 — Cys296 | 16.71 | нет связи |
Все четыре цистеина являются свободными. Это типично для внутриклеточных белков анаэробных термофилов, где окислительная среда, необходимая для образования S–S связей, отсутствует.
Стекинг — нековалентное взаимодействие между π-электронными системами ароматических колец (Phe, Tyr, Trp, His). Различают параллельный (face-to-face, угол нормалей 0–30°) и T-образный (edge-to-face, угол 60–90°) стекинг. Расстояние центроид–центроид при стекинге ≤ 5.5 Å.
В структуре 2X42 содержится 76 ароматических остатков. Поиск выявил 14 пар с расстоянием центроид–центроид ≤ 5.5 Å. Два типа стекинга задокументированы ниже.
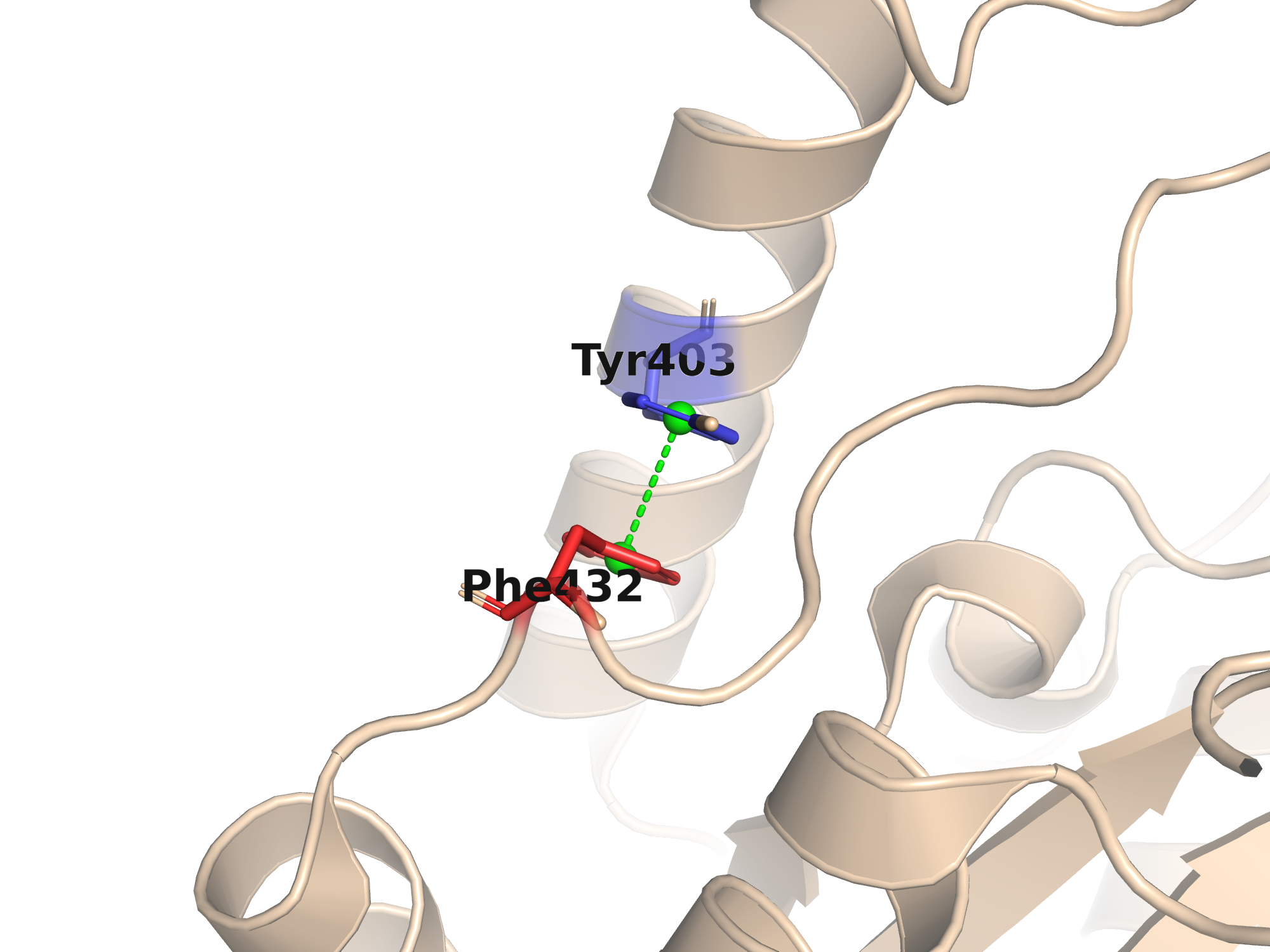


| Пара | d, Å | Угол | Тип стекинга |
|---|---|---|---|
| Tyr403 — Phe432 | 3.78 | 5.3° | Face-to-face (параллельный) |
| Tyr580 — Phe594 | 4.47 | 17.5° | Face-to-face (параллельный) |
| Phe469 — Tyr470 | 4.98 | 83.2° | T-образный |
| Tyr582 — Tyr583 | 5.39 | 76.7° | T-образный |
В структуре PDB 2X42 проанализированы пять типов нековалентных взаимодействий: