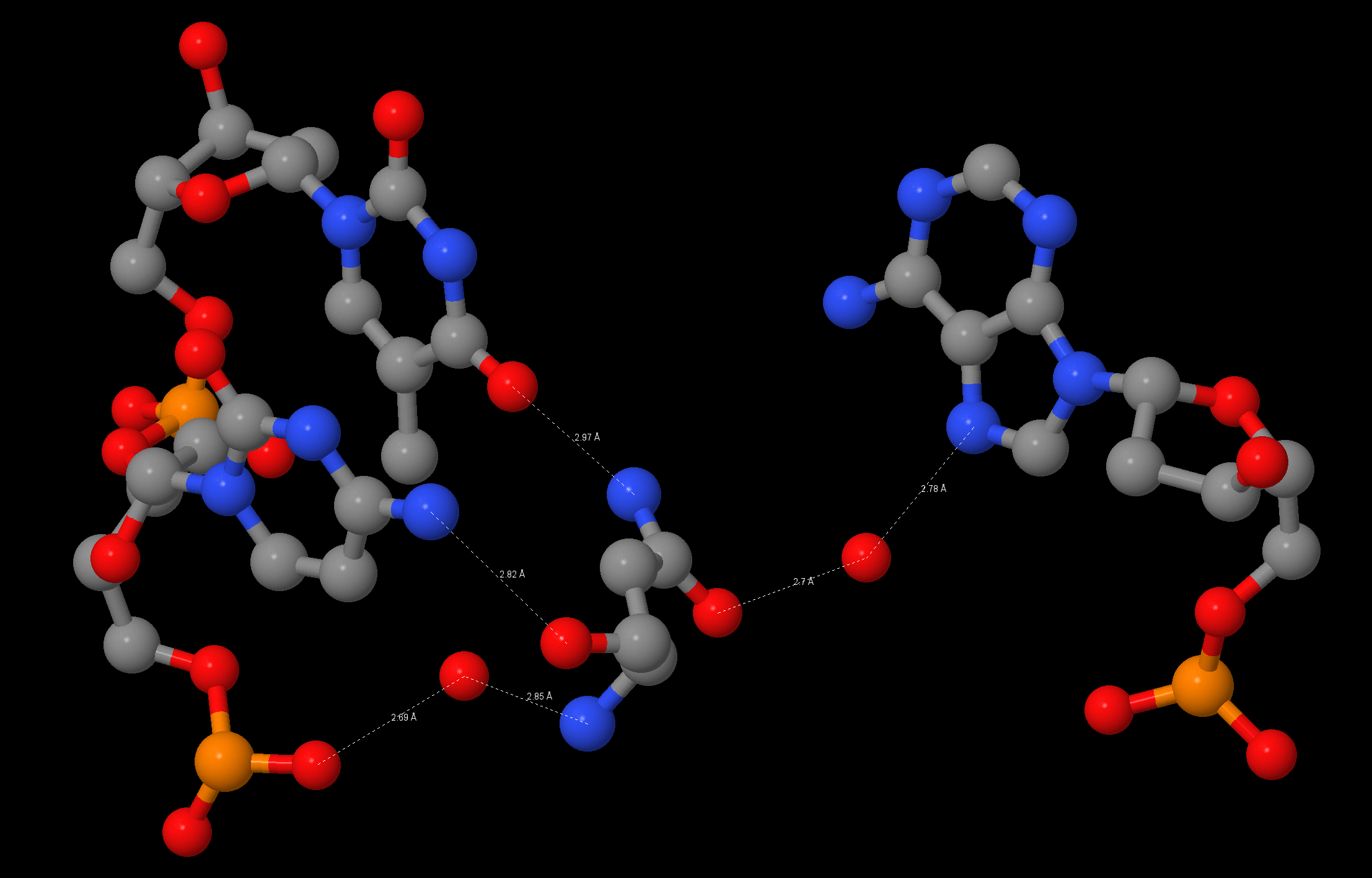
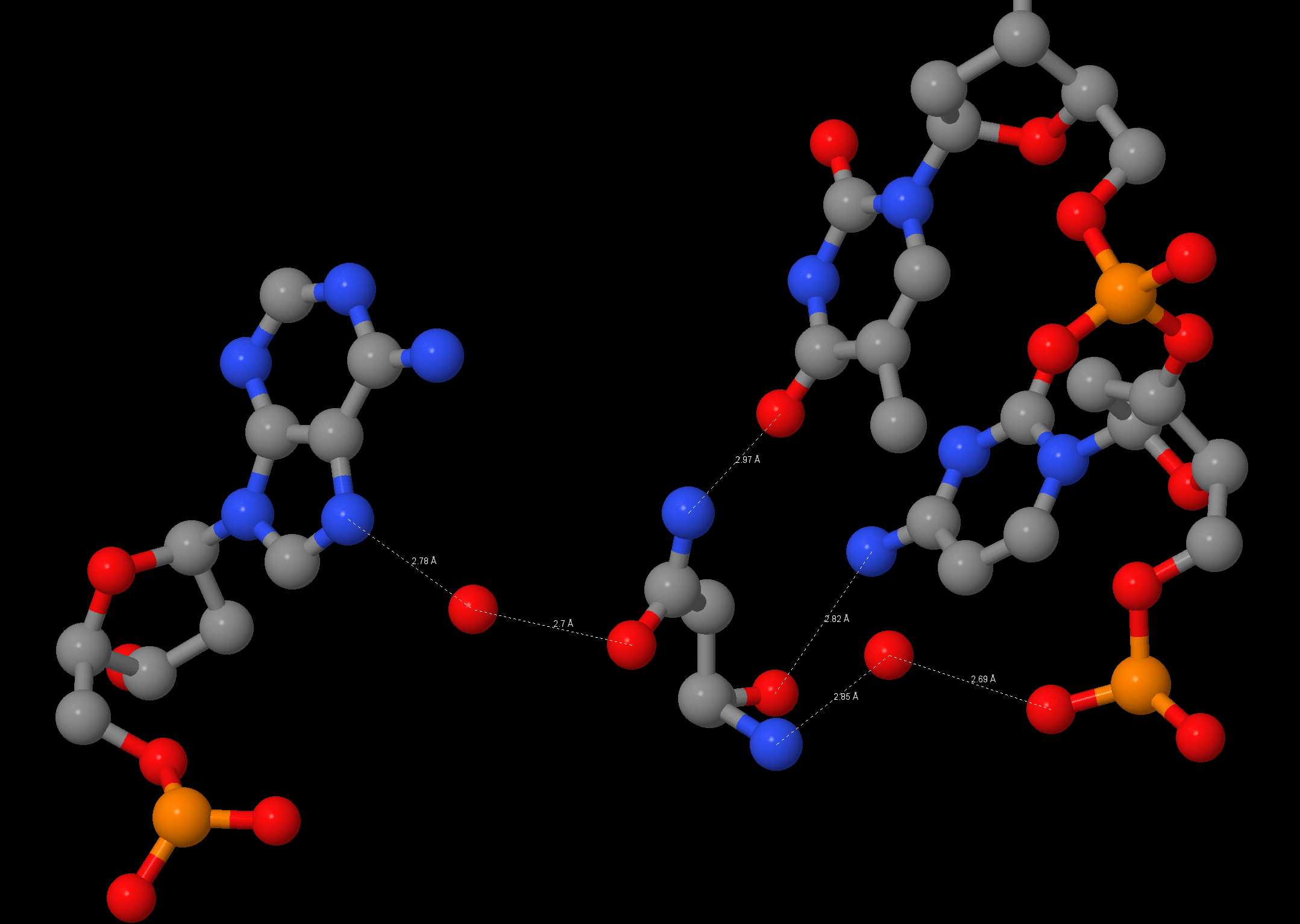
Изображение водородных связей между атомами [ASN]140:B и атомами ДНК: [ASN]140:B.OD1-[HOH]833:B.O-[DA]5:C.N7 [ASN]140:B.ND2-[DT]28:D.O4 [ASN]140:B.O-[DC]27:D.N4 [ASN]140:B.N-[HOH]33:D.O-[DC]27:D.OP2
| Участок структуры | Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
|---|---|---|---|
| Акцепторный стебель | 5' 501-572 3' 5' 502-571 3' 5' 503-570 3' 5' 504-569 3' 5' 505-568 3' 5' 506-567 3' 5' 507-566 3' 7 пар | предсказано 5 пар из 7 реальных | предсказано 7 из 7 |
| D-стебель | 5' 510-525 3' 5' 511-524 3' 5' 512-523 3' 5' 513-522 3' 4 пары | ничего не предсказано | предсказано 4 из 4 |
| T-стебель | 5' 549-565 3' 5' 550-564 3' 5' 551-563 3' 5' 552-562 3' 5' 553-561 3' 5 пар | ничего не предсказано | предсказано 5 из 5 |
| Антикодоновый стебель | 5' 538-532 3' 5' 539-531 3' 5' 540-530 3' 5' 541-529 3' 5' 542-528 3' 5' 543-527 3' 5' 544-526 3' 7 пар | ничего не предсказано | предсказано 5 из 7 |
| Общее число канонических пар нуклеотидов | 19 | 5 | 20 |
2CV0C: Score 15: 5/5 (100%) matches, 0 gaps
3 cccca 7
|||||
69 ggggt 65
|
Только уменьшив значение параметра "Minimum score threshold" программы einverted до 10, удалось найти инвертированные участки в данной мне нуклеотидной последовательности. Как выяснилось, найденный сегмент вторичной структуры тРНК соответствует акцепторному стеблю. Стоит заметить, что такой участок оказался единственным выданным программой einverted. Из этого, а так же из того факта, что вообще этот участок удалось найти только понизив параметр "Minimum score threshold" до 10, можно заключить, что структура данной тРНК целиком зависит от белка, с которым образует комплекс. Это объясняется тем, что входным файлом программы find_pair является файл формата PDB (2cv0_old.pdb), где включены данные о тРНК в комплексе с белком, а программа einverted на вход принимает файл формата FASTA (2cv0c.fasta), в котором находится информация только о тРНК. То есть find_pair учитывает влияние белок-тРНК взаимодействий на вторичную структуру тРНК, а einverted - нет.
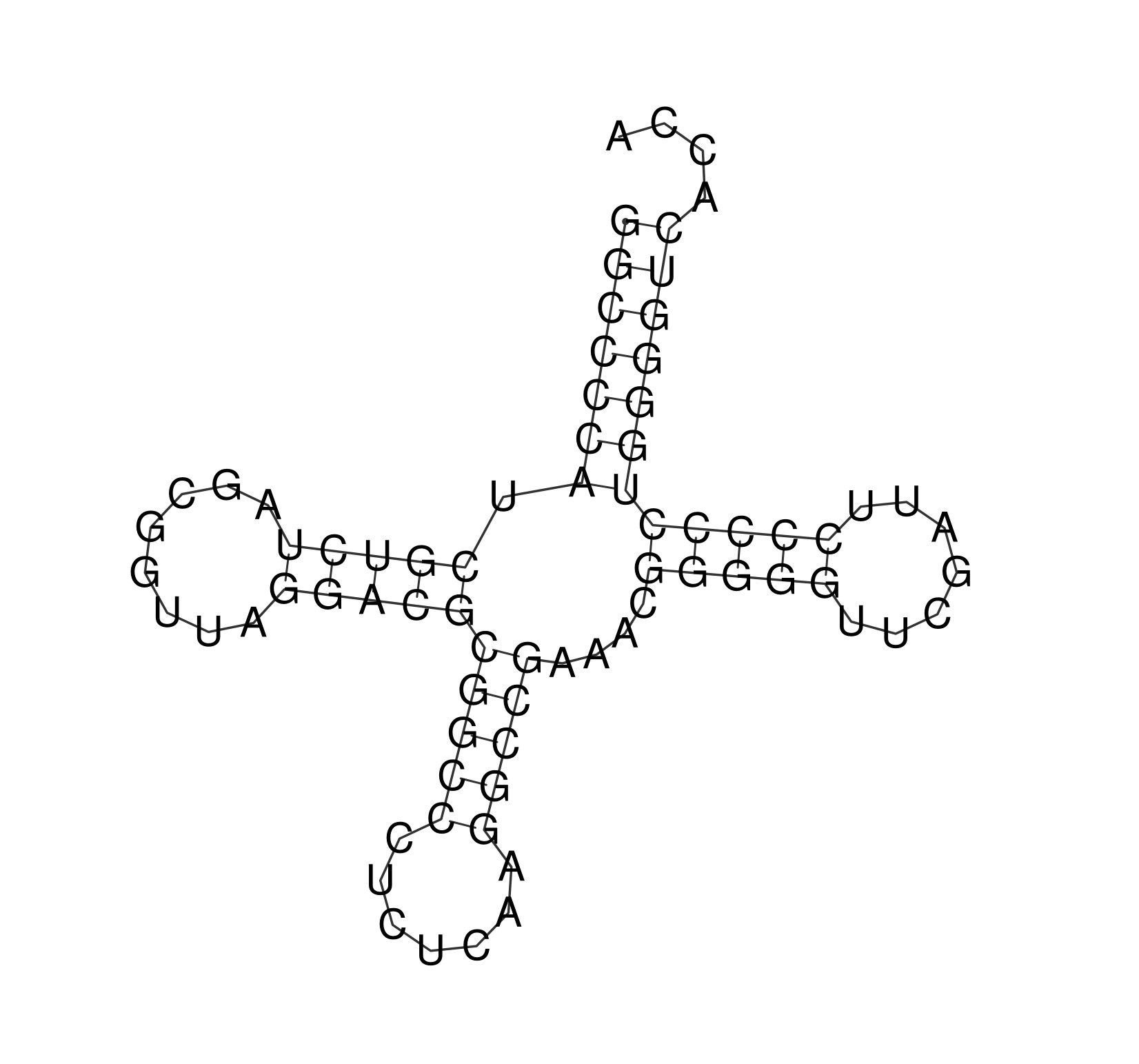
Предположительная вторичная структура тРНК, созданная по алгоритму Зукера, приведена на изображении ниже. Она в основном соответствует данным, выданными программой find-pair (несовпадения опять же, скорее всего, связаны с тем, что тРНК связан с белком), и была получена с параметрами по умолчанию (то есть с первого запуска программы RNAfold).
Скрипт-файл с определениями множеств set1, set2, set3
| Контакты атомов белка с | Полярные | Неполярные | Всего |
|---|---|---|---|
| остатками 2'-дезоксирибозы | 7 | 45 | 52 |
| остатками фосфорной кислоты | 17 | 14 | 31 |
| остатками азотистых оснований со стороны большой бороздки | 11 | 22 | 33 |
| остатками азотистых оснований со стороны малой бороздки | 5 | 0 | 5 |
Как видно из таблицы, малая бороздка мало задействована в ДНК-белковых контактах (это можно объяснить тем, что нахождение в области, где атомы азотистых оснований сближены, энергетически невыгодно для атомов белка). Выяснилось, что поддержание комплекса обеспечивается в основном контактами атомов белка с атомами фосфорных остатков и 2'-дезоксирибозы (образовывая связи непосредственно или же чере молекулу воды как посредника); это закономерно, так как пространственно атомы сахаро-фосфатного остова более доступны для образования связи, чем атомы азотистых оснований, которые направлены вглубь молекулы. Если говорить о доле участия полярных и неполярных контактов, то, возможно, это зависит от количества атомов соответствующего типа в остатке. Так, в молекуле дезоксирибозы на 3 полярных атомов кислорода приходится 5 неполярных углерода, и, соответсвенно, неполярных контактов с атомами белка дезоксирибоза образует больше, чем полярных. Если же рассматривать остатки фосфорной кислоты, то в них на 1 неполярный фосфор приходится 2 полярных кислорода, и полярных контактов с атомами белка они образуют больше. Для остатков азотистых оснований со стороны малой бороздки такая закономерность не наблюдается: несмотря на то, что число полярных и неполярных атомов, обращенных в сторону большой бороздки, примерно одинаково, неполяных контактов с атомами белка они образуют в 2 раза больше, чем полярных.
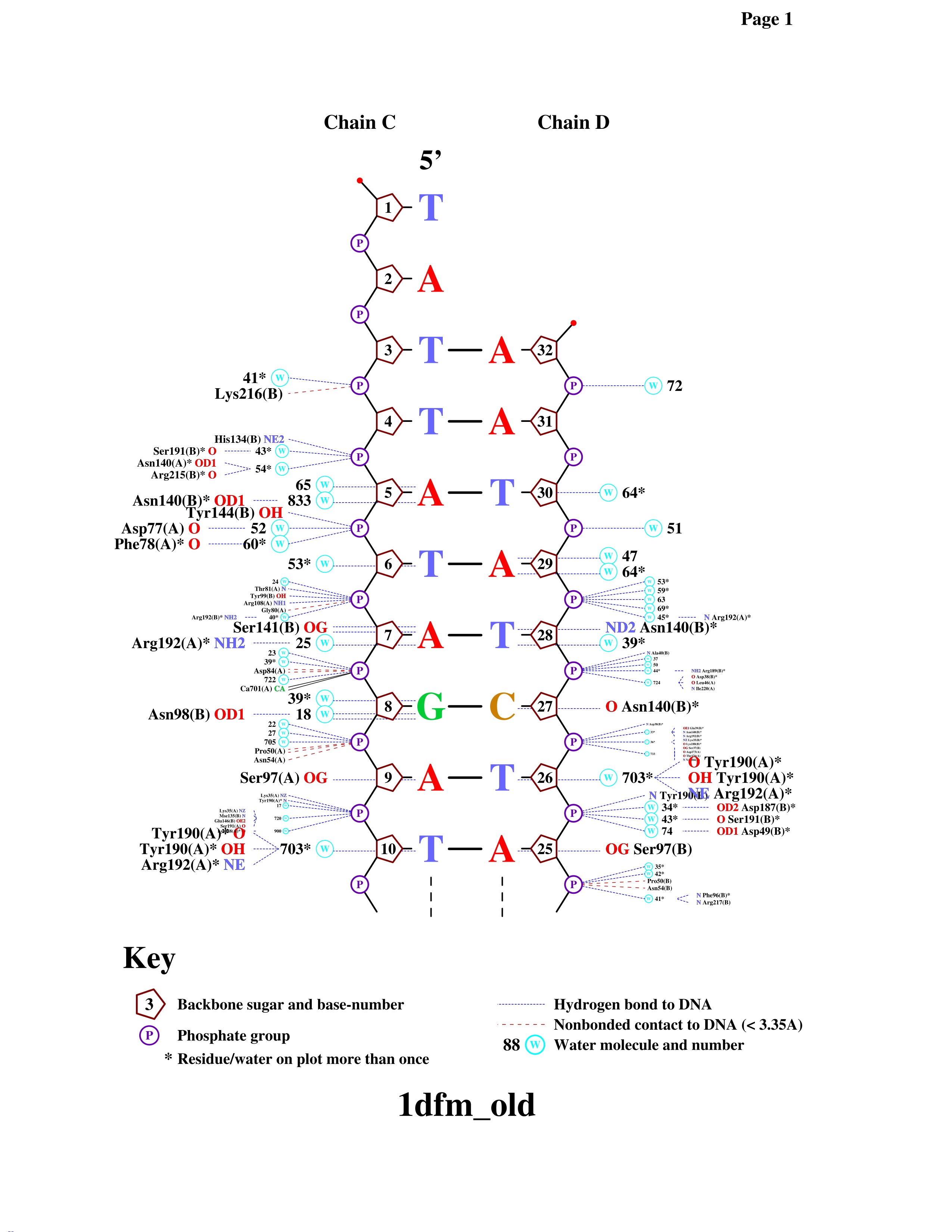
Наибольшее количество контактов с ДНК имеет Tyr190(A): 1 непосредственный контакт - водородная связь, и 4 через посредника - воду(703), тоже водородные связи.
Как мне кажется, Asn140(B) - наиболее значимый аминокислотный остаток для распознования последовательности ДНК, так как он имеет наибольшее количество контактов (3) с различными азотистыми основаниями (причем находящимися на разных цепях) и к тому же одну связь с фосфатом. Если говорить конкретно, то атом OD1 аспарагина образует водородную связь с N7 аденина через связь с водой, ND2 аспарагина - с O4 тимина, O аспарагина - с N4 цитозина и N аспарагина образует водородную связь с OP2 фосфата через молекулу воды.
Изображение водородных связей между атомами [ASN]140:B и атомами ДНК: [ASN]140:B.OD1-[HOH]833:B.O-[DA]5:C.N7 [ASN]140:B.ND2-[DT]28:D.O4 [ASN]140:B.O-[DC]27:D.N4 [ASN]140:B.N-[HOH]33:D.O-[DC]27:D.OP2
© Агаева Зара, 2018