В данном задании требуется сравнить две модели структур фермента Notum, который является карбоксилэстеразой.
Каждая из моделей была получена методом рентгеноструктурного анализа с последующим уточнением структуры.
Эксперимент для 7B2Y был проведен в 2020 году, из которого была получена структура с разрешением 1.23 Å
на основании 97995 рефлексов. При этом доля числа рефлексов от теоретически возможного числа рефлексов в диапазоне
разрешений от лучшего к худшему = 100%. Температура проведения диффракционнго эксперимента 100 К. Длина волны падающего
излучения 0.9763 Å. Так же здесь был указан метод уточнения структуры: молекулярное замещение.
Эксперимент для 4UYZ был проведен в 2012 году, из которого была получена структура с разрешением 2.8 Å
на основании 38300 рефлексов. При этом доля числа рефлексов от теоретически возможного числа рефлексов в диапазоне
разрешений от лучшего к худшему = 89.1%. Температура проведения диффракционнго эксперимента 100 К. Длина волны падающего
излучения почему-то не указана.
Также было интересно посмотреть на параметры элементарных ячеек кристаллов белков.
Таблица 1. Параметры элементарной ячейки для 7B2Y.
Длина (Å)
Угол (°)
a = 60.01
α = 90
b = 71.54
β = 90
c = 78.02
γ = 90
Таблица 2. Параметры элементарной ячейки для 4UYZ.
Длина (Å)
Угол (°)
a = 60.829
α = 90
b = 193.88
β = 91.88
c = 75.717
γ = 90
После открытия структур для обоих ID, оказалось, что в элементарную ячейку 7B2Y входит один белок, а в
элементарную ячейку 4UYZ четыре белка. Для дальнейшей работы я выделил лишь один белок из 4UYZ.
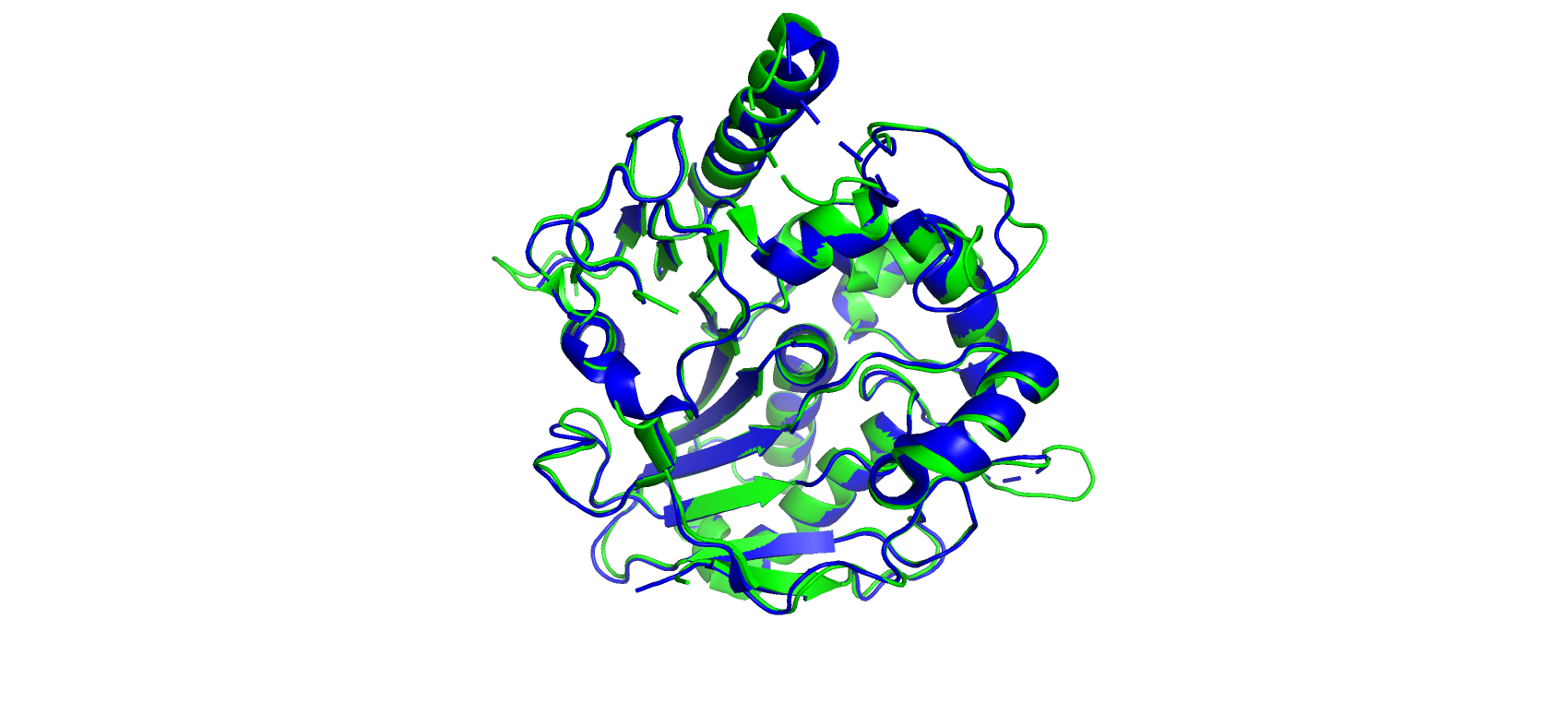
Чтобы наглядно продемонстрировать и для большего удобства поиска различий двух структур струтура одного выбранного белка
из 4UYZ была выравнена по структуре белка 7BYZ с помощью команды super.
Рис 1. Выровненные структуры 7BYZ (зеленый) и 4UYZ (синий)
Прежде всего выделяются участки с гэпами (Рис. 1). При этом в модели 7BYZ этих пропусков меньше (она, кстати, по описанию имеет
лучшее разрешение). Видимо после эксперимента не хватило информации для предсказания электронных областей в подобных участках.
Рис 2. Участки с пропусками в структурах 7BYZ (зеленый) и 4UYZ (синий) 348-358 аминокислоты.
Помимо этого видно, что модели не идеально похожи друг на друга как в ходе остова: спирали имеют местами разную структуру (Рис. 3);
некоторые петли имеют разную ориентацию (Рис. 4). Так и в ориентации боковых цепей аминокислот (Рис. 5).
Рис 3. Разный ход спиралей в структурах 7BYZ (зеленый) и 4UYZ (синий) 303-315 аминокислоты.
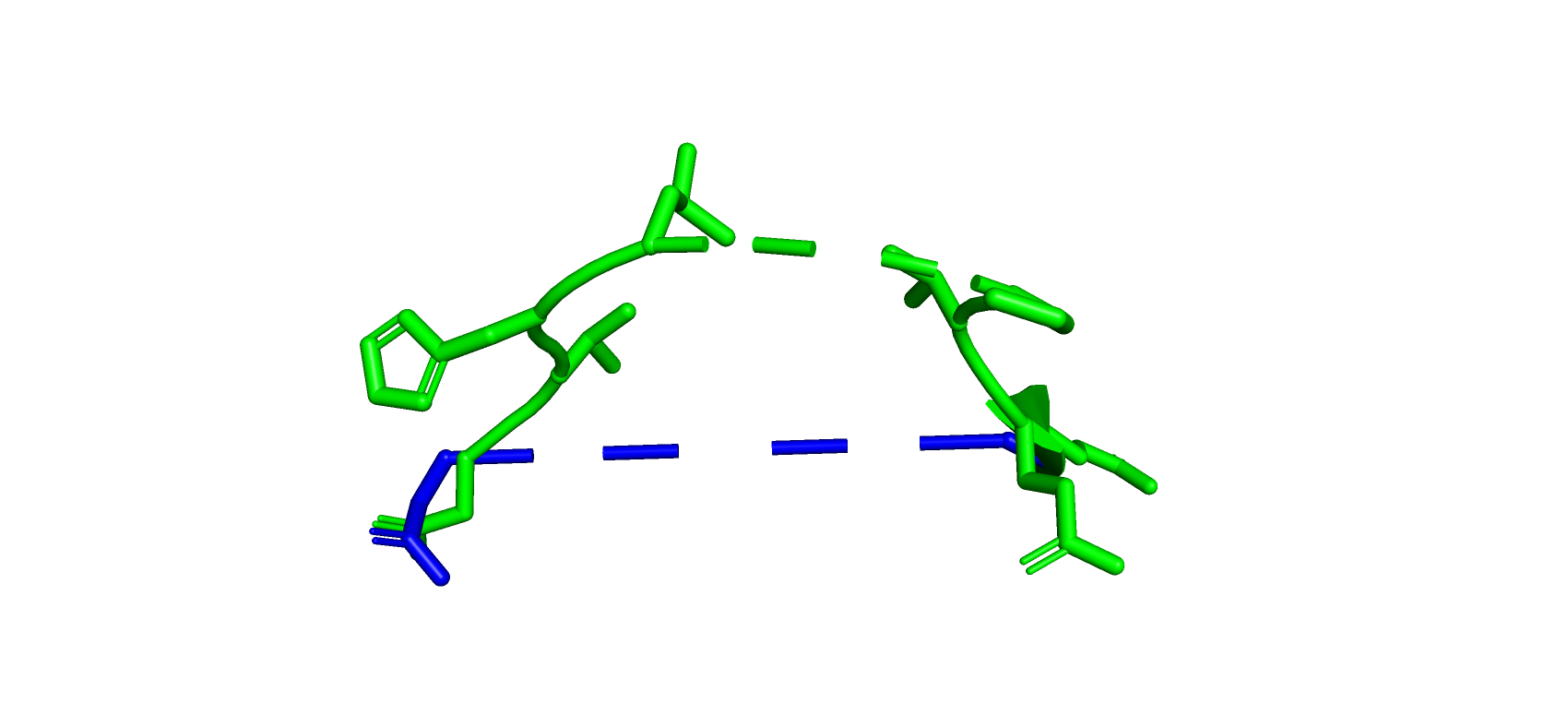
Рис 4. Разные ориентации петель в структурах 7BYZ (зеленый) и 4UYZ (синий) 265-286 аминокислоты.
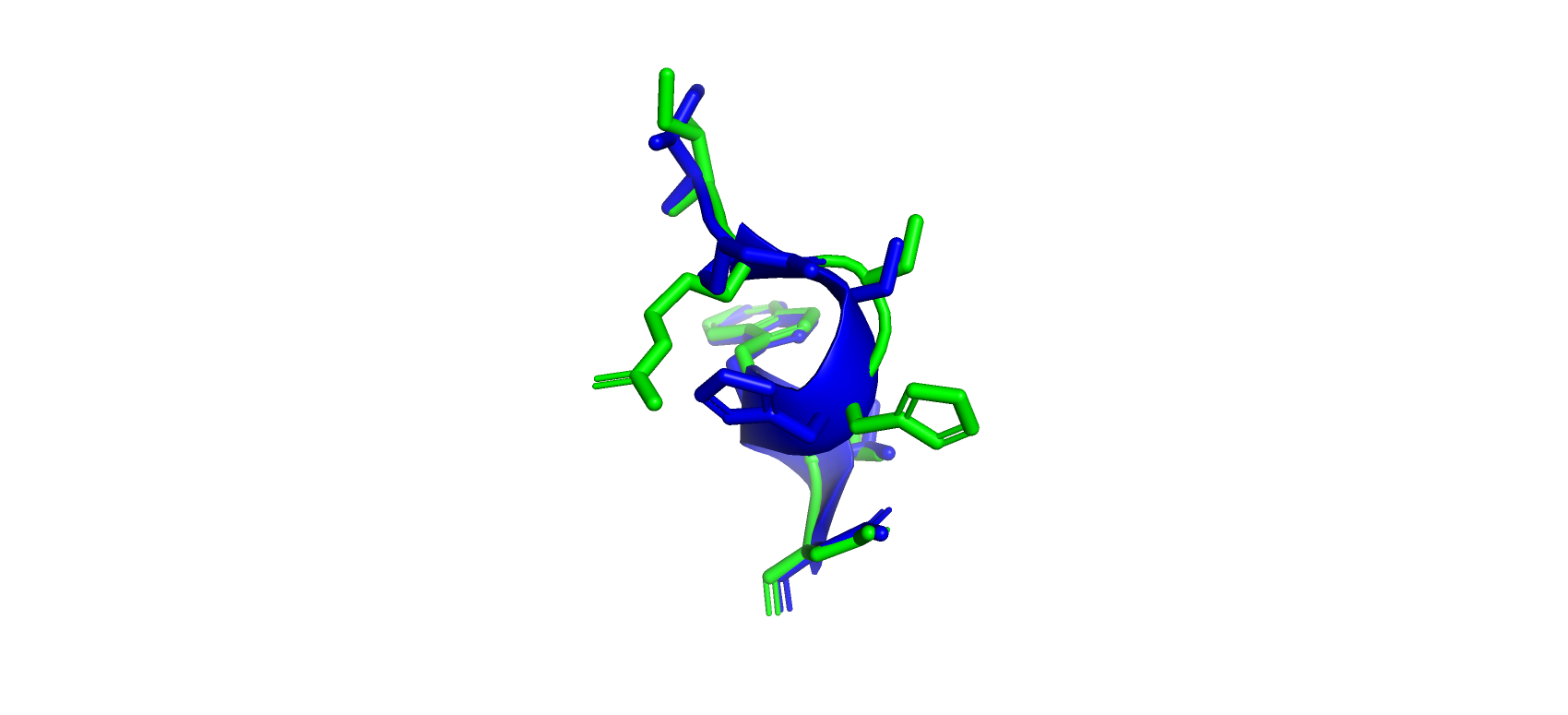
Рис 5. Демонстрация как совпадающих ориентаций боковых радикалов, так и различных в структурах 7BYZ (зеленый) и 4UYZ (синий) 393-399 аминокислоты.
Далее с помощью команды isomesh были изучены карты электронной плотности с уровнем подрезки 2 двух структур для их
сравнения.
Для этого были взяты одинаковые участки белковой последовательности с Gln 258 по Asp 264 в обеих структурах.
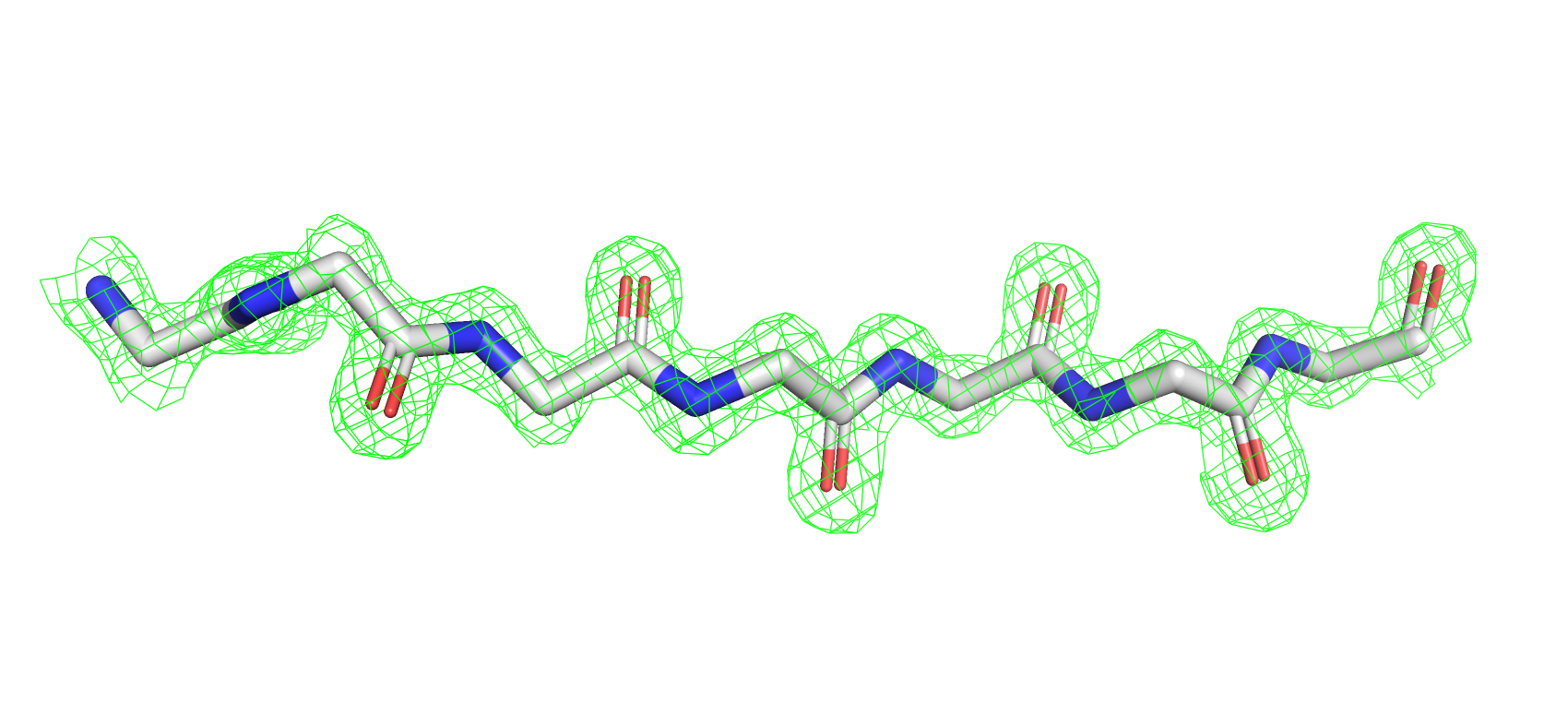
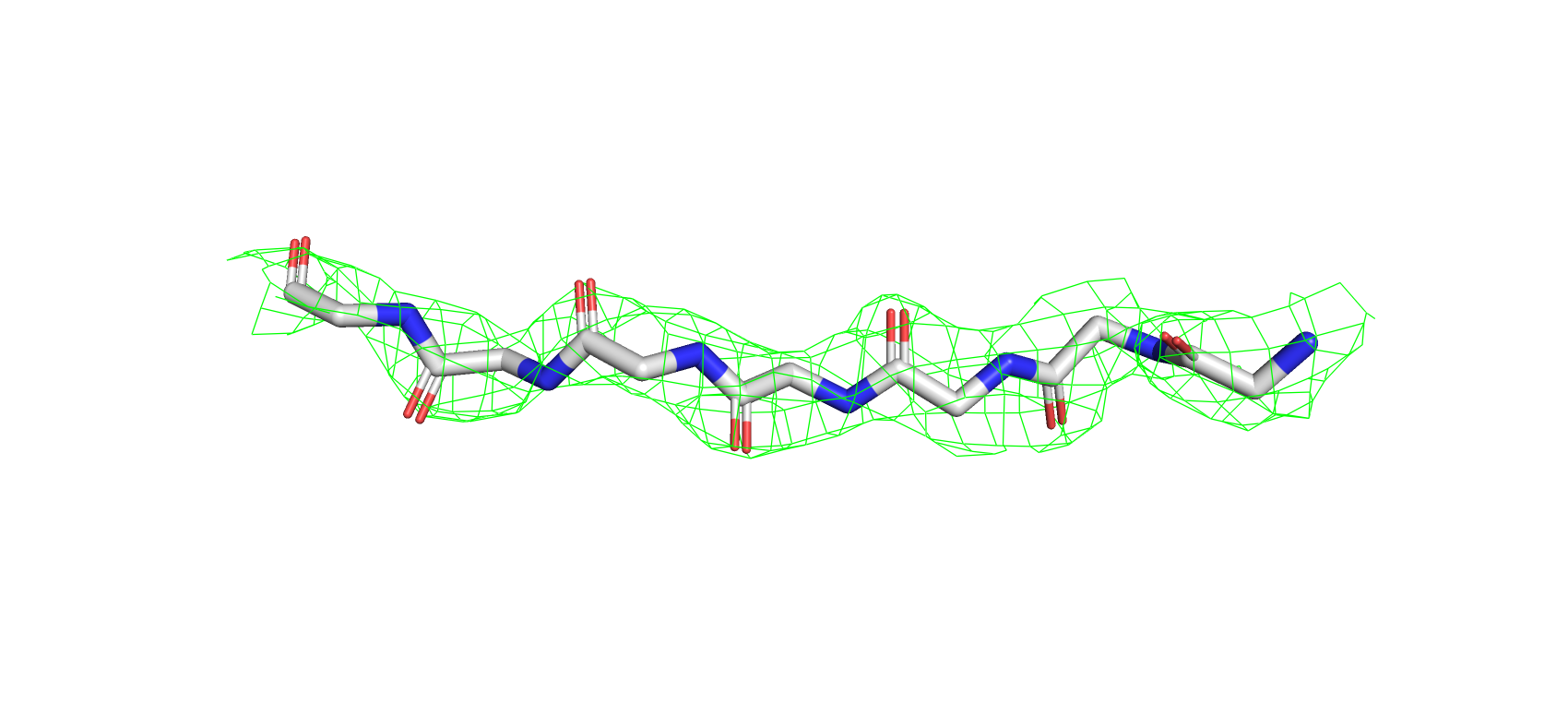
На рис. 6 видно, что для данного участка структуры 7BYZ электронная плотность формирует вокруг атомов остова
сферическую поверхность, что соответствует теоретическим представлениям о распределении ЭП.
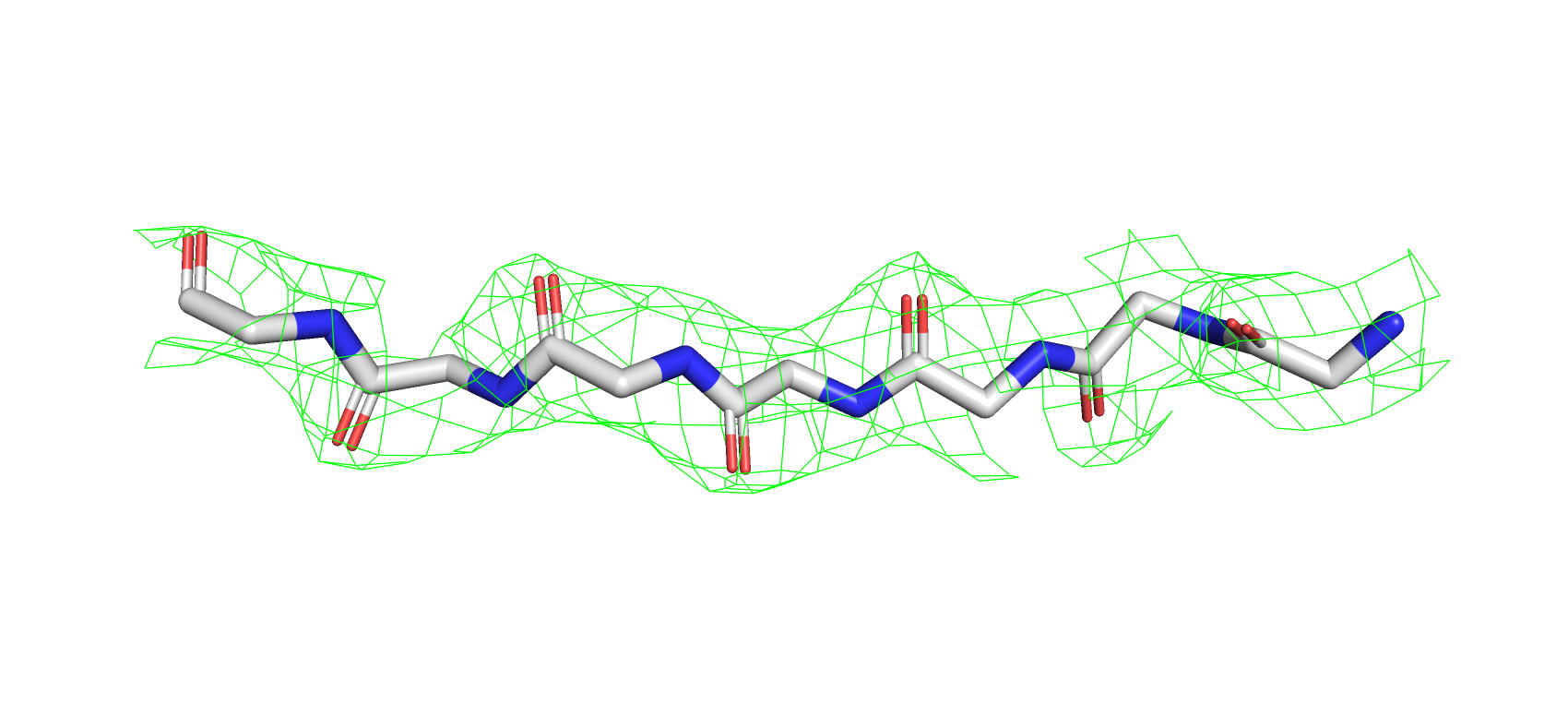
Электронная плотность на этом же участке 4UYZ имеет довольно крывую форму (рис.7), не очерчивая отдельные атомы остова
как в предыдущей структуре. Более того есть места где, например, расположение атома в структуре вылазеет за пределы
электронной плотности (по крайней мере при подрезке 2). Подрезка уровня 1 решает эту проблему, однако, появляются участки с
довольно большими дырками в электронных плотностях (рис. 8). При этом игра с curve не дала изменений, т.к. при больших значениях
появлялиь электронные плотности в других участках структуры.
Кажется, что структура 4UYZ имеет худшее разрешение, что согласуется с информацией из записей PDB. Связано это может быть в
в первую очередь из-за меньшего числа рефлексов в эксперименте, меньшей доле числа рефлексов от теоретически возможного
числа рефлексов в диапазоне разрешений от лучшего к худшему и инструментальные погрешности (чувствительность приборов, длина
излучаемой волны).
Рис 6. Электроннная плотность фрагмента 7BYZ на уровне подрезки 2, carve = 1.
Рис 7. Электроннная плотность фрагмента 4UYZ на уровне подрезки 2, carve = 1.6.
Рис 8. Электроннная плотность фрагмента 4UYZ на уровне подрезки 1, carve = 1.6.
Задание 2
Для визуализации электронной плотности на разных уровнях подрезки я использовал структуру 7KEQ.
Для сравнения разных уровней подрезки электронной плотности были построены три рисунка с уровнями подрезки от 1 до 3.
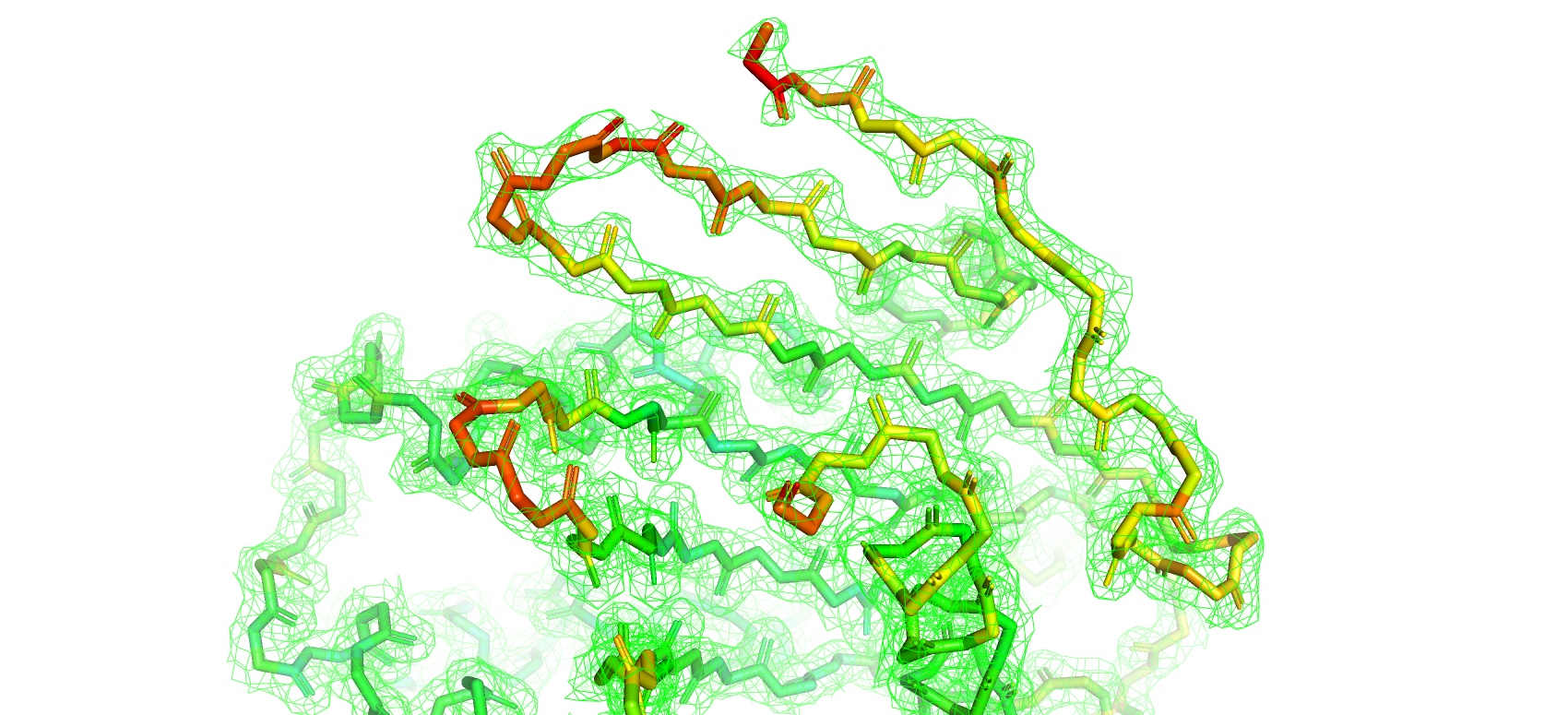
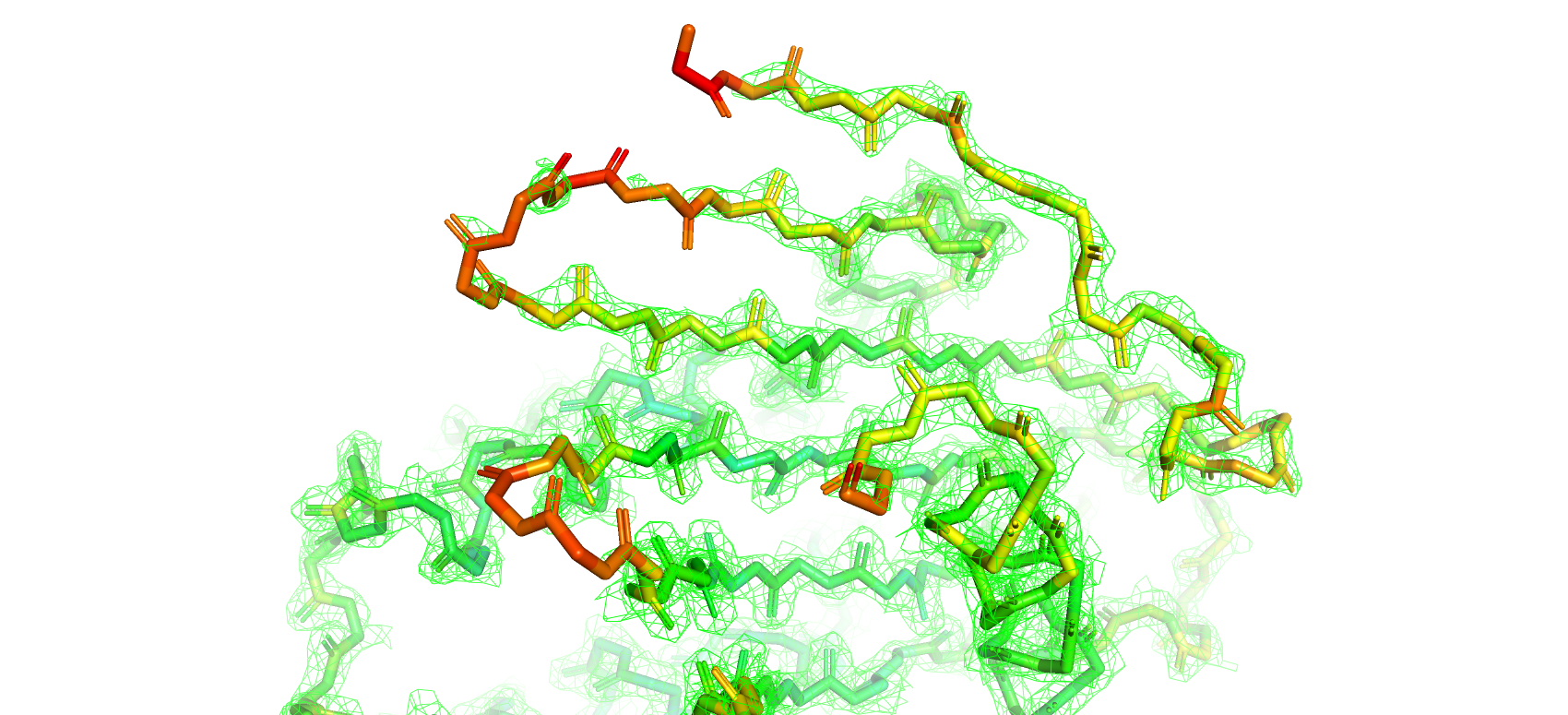
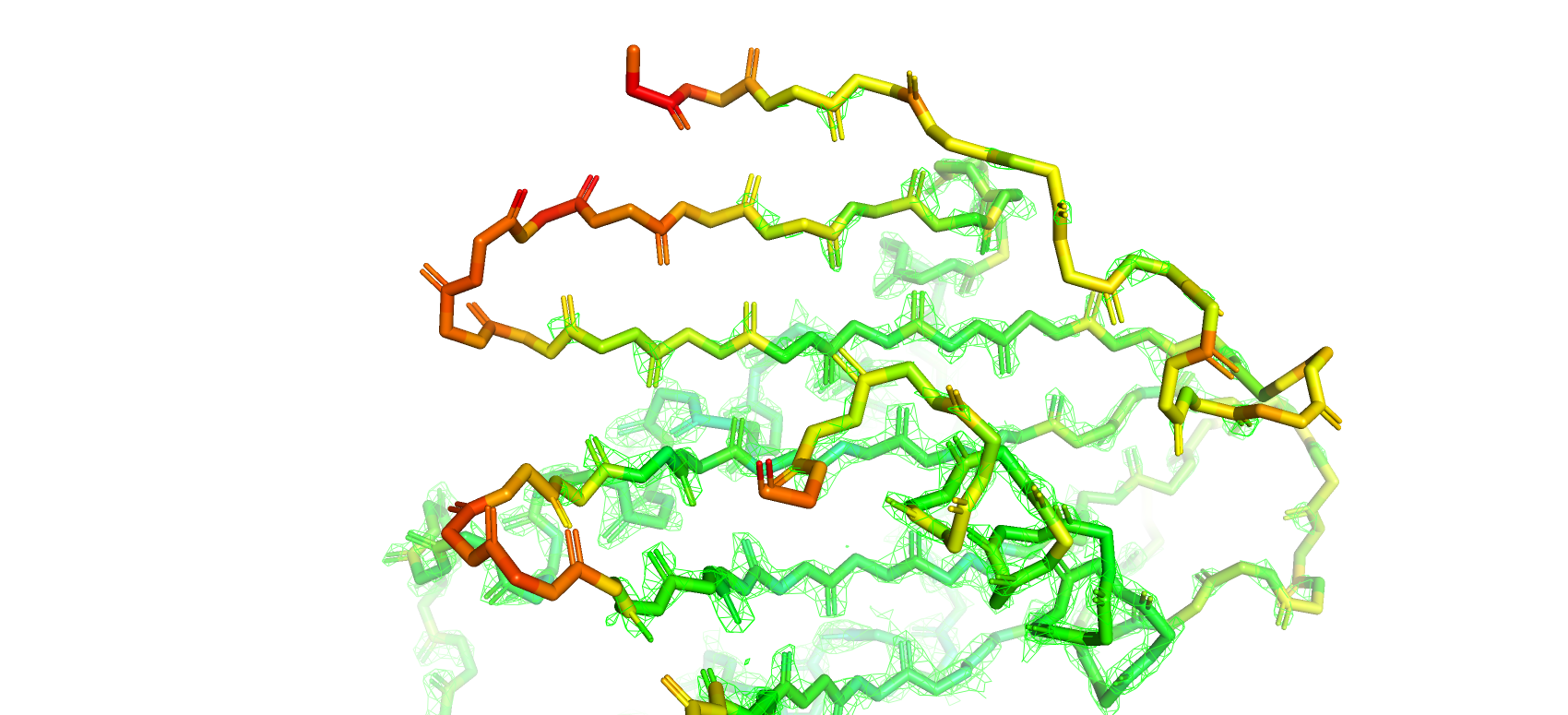
Для удобства анализа структура была покрашена по B-фактору, который является мерой отклонения позиции атома от его среднего положения
по всем ячейкам кристалла (от синего - самого маленького до красного - самого большого значения B-фактора)
Далее были построены электронные плотности вокруг атомов остова при разных уровнях подрезки 1, 2, 3 (рис. 9, 10, 11 соответственно)
Как и следовало ожидать при увилечении уровня подрезки плотность отображается все на меньшем и меньшем количестве атомов
остова структуры. Особенно сильно это касается участков с высокими значениями B-фактора (желтый-красный цвета).
В моем случае наибольшие значения B-фактора и худшие ЭП на каждом уровне подрезки наблюдается у C- и N-концов
белка, а также в некоторых поворотах с одного β-листа на другой. Очевидно, что из-за высокой подвижности атомов расположенных
в этих участках ЭП (мало удерживающих сил: взаимодействий с окружающими остатками или поверхностное расположение участка)
размывается по большему объему из-за чего при увеличении подрезки она значительно уменьшается. А участки с наименьшей подвижностью
иногда даже имеют при подрезке 3 довольно качественные (близкие к сферическим) поверхности ЭП.
Рис 9. Электроннная плотность остова структуры 7KEQ на уровне подрезки 1, carve = 1.5.
Рис 10. Электроннная плотность остова структуры 7KEQ на уровне подрезки 2, carve = 1.5.
Рис 11. Электроннная плотность остова структуры 7KEQ на уровне подрезки 3, carve = 1.5.
Задание 3
В этом задании были построены электронные плотности вокруг лиганда, который присутствует в этой модели, также на
уровнях подрезки 1, 2 и 3 (рис. 12, 13, 14 соответственно).
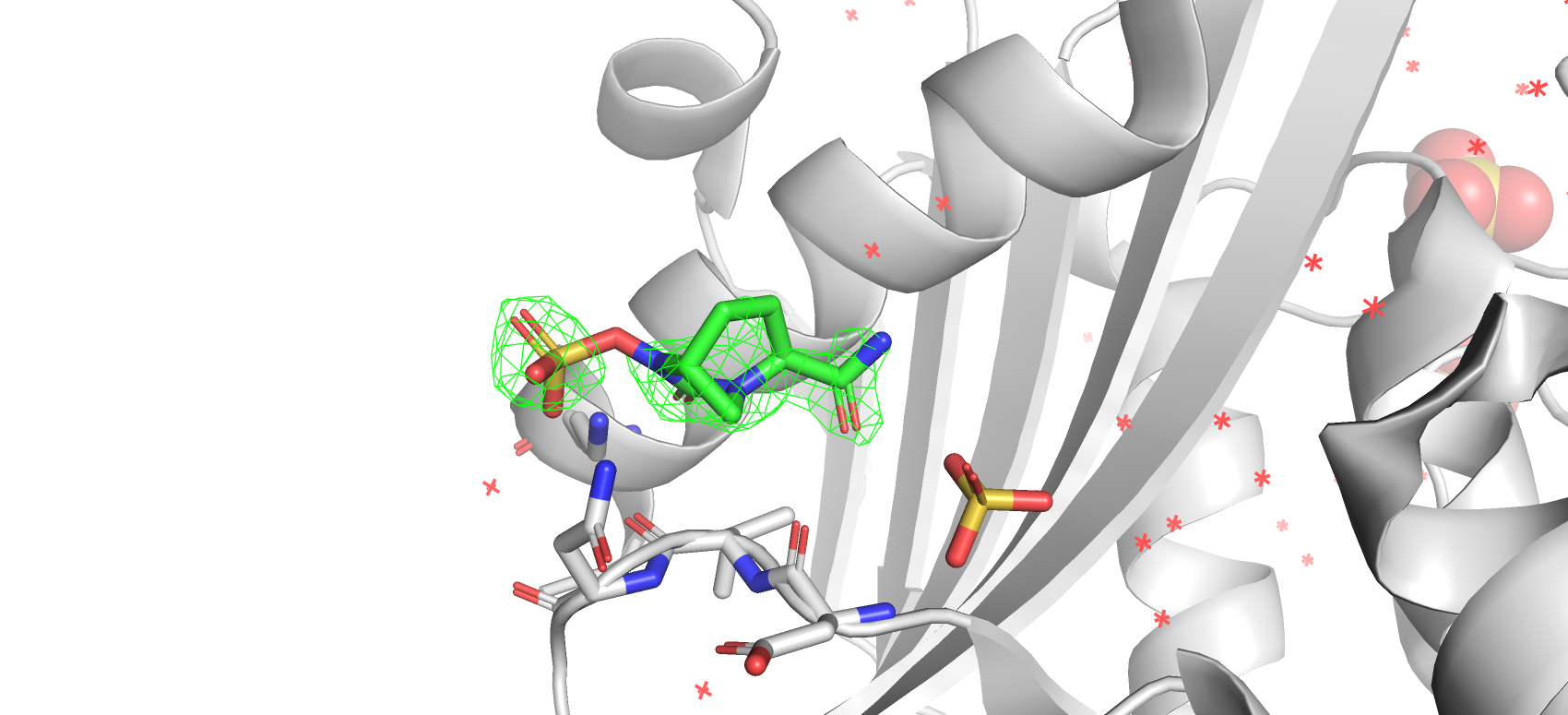
В моей структуре несколько лигандов, однако, мой взгляд упал на интересный по своей структуру Avibactam (c бициклом).
На уровне подрезки 1 ЭП располагалась на сульфогруппе, на бицикле (на кислороде кетогруппы включительно) и на амидной группе.
Единственным в сторонке остался атом кислорода между сульфогруппой и бициклом, и один из фтомов углерода в бицикле.
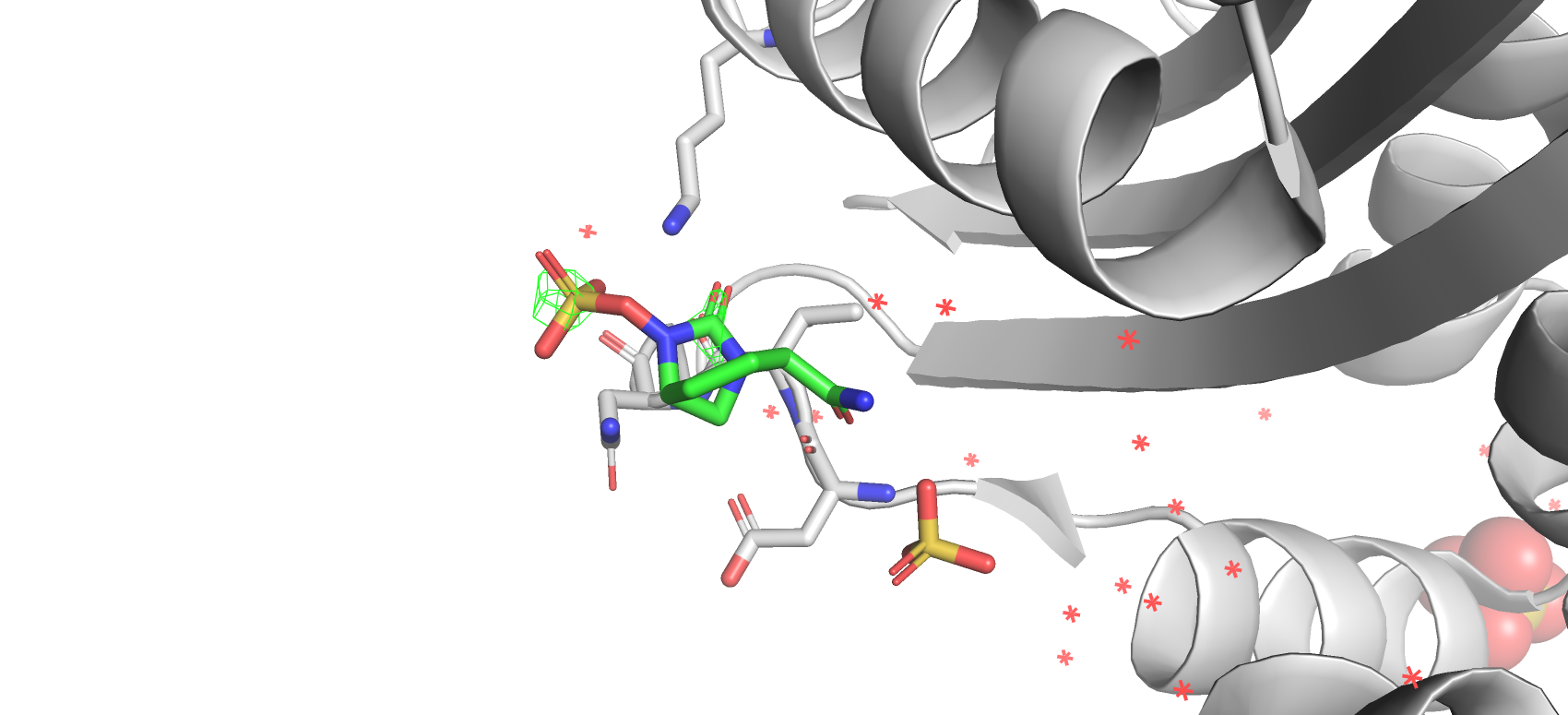
На уровне подрезки 2 ЭП осталась лишь вокруг атома серы в сульфогруппе и в области карбоксильной группы в бицикле.
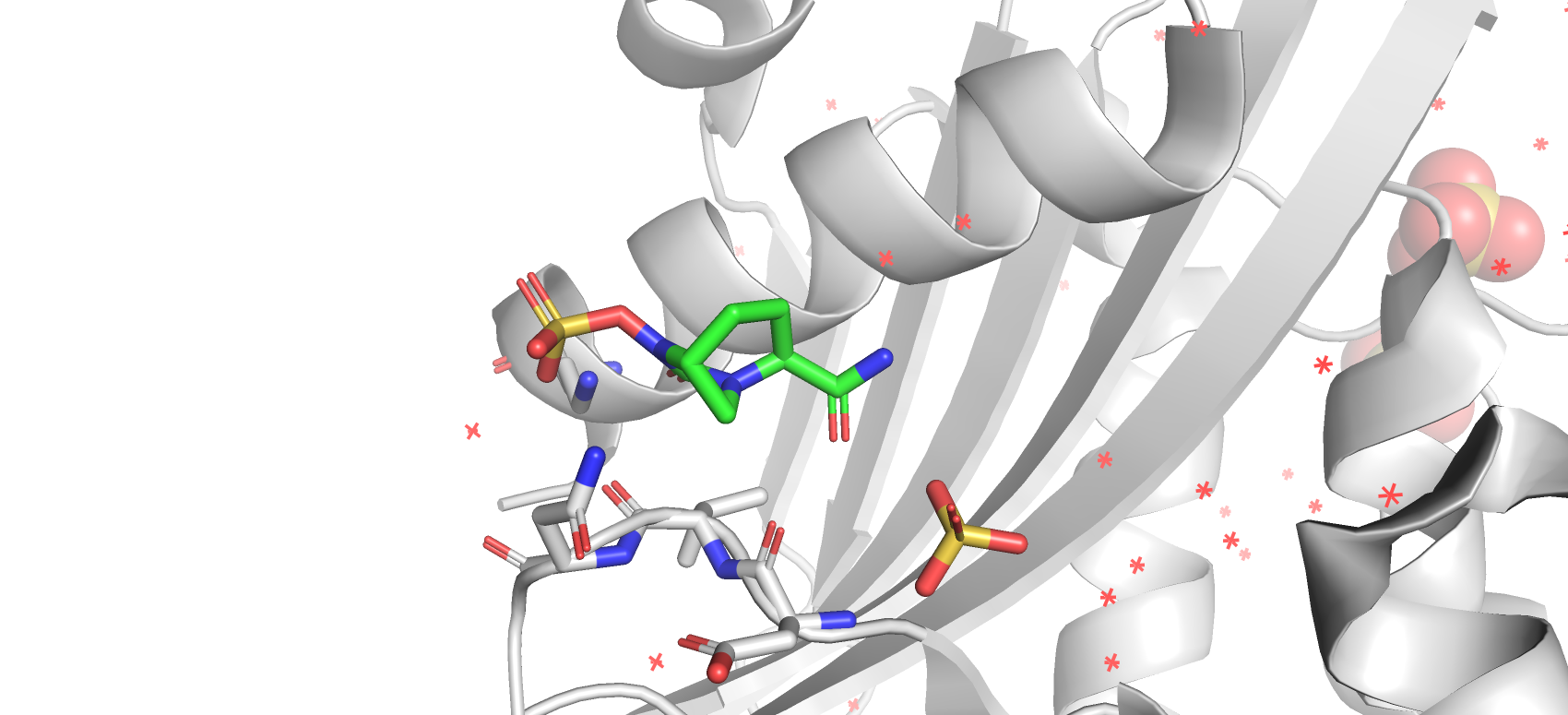
На уровне подрезки 3 ЭП вообще не располагалась вокруг какого-либо атома лиганда.
Можно отметить, что с повышением уровня подрезки ЭП остается в облостях с наибольшей ЭО. Почему при подрезке 2 ЭП
остается вокруг атома серы, а не вокруг атомов кислорода интересный вопрос. А вот куда деваются ЭП при подрезке 3 еще более интересный вопрос (вероятно сам лиганд там удерживается слабо и не во всех ячейках кристала находился в сайте связывания).
Вероятно это все таки связано с недостаточным количеством информации для именно ЭП лиганда - ЭП впринципе недостаточно хорошо реконструирована для этого лиганда (хотя разрешение самой структуры не такое уж и плохое 2 Å)
Рис 12. Электроннная плотность лиганда структуры 7KEQ на уровне подрезки 1, carve = 1.8.
Рис 13. Электроннная плотность остова структуры 7KEQ на уровне подрезки 2, carve = 1.8.
Рис 14. Электроннная плотность остова структуры 7KEQ на уровне подрезки 3, carve = 1.8.