Практикум #9 Структура/функция. Поиск по структуре
Введение
Мой Uniprot ID: A0A6Q2YM18.
Этому идентификатору соответствует белок с названием: субъединица 1 рецептора гамма-аминомасляной кислоты типа B из
организма Esox lucius (щука обыкновенная).
Чтобы разобраться в механизме работы этого рецептора и его устройстве я решил поискать какую-нибудь не очень старую
статью. В итоге мне удалось ее найти: doi: 10.3390/molecules25133093. Журнал средненький (impact factor 4.4), зато
статья 2020 года и содержит в себе упоминания основных структурных особенностей, связанных с рецепторной функцией,
устройством полноценной структуры рецептора, аллостерической регуляцией, которые я опишу ниже.
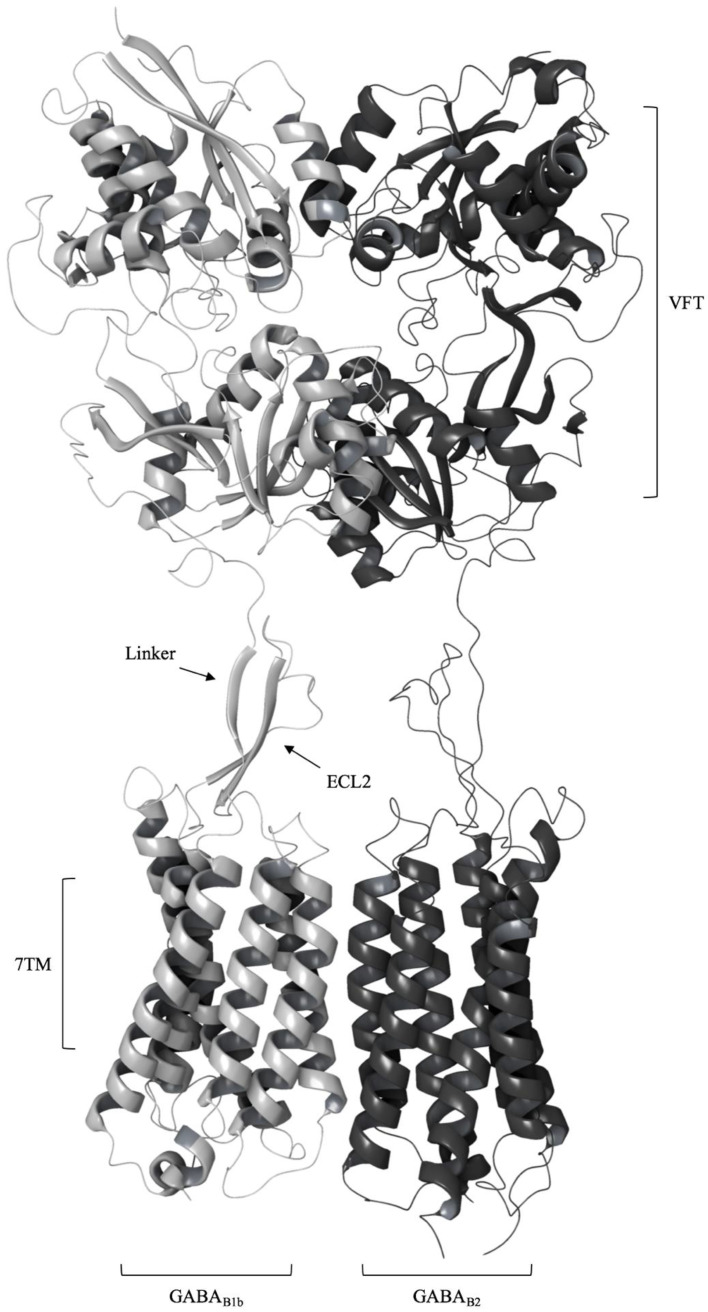
GABAB-R является гетеродимером, который состоит из двух субъединиц: B1 (которая является целью этого задания) и B2.
Каждая субъединица состоит из внеклеточного домена (VFT), трансмембранного домена, состоящего из 7 спиралей, и линкера
между ними (рис. 1). При изучении РСА структуры VFT было отмечено что B1 субъединица имеет сайт связывания (ортостерический),
а для B2 не было показано взаимодействие с лигандами. Однако для B2 субъединицы указано основное участие в рекрутировании
G-белка (взаимодействие через его 7TM субъединицу) и наличие аллостерического сайта связывания.
Рис 1. Схематическая репрезентация GABAB-R, взятая из представленной выше статьи. VFT -
внеклеточный домен, 7TM - трансмембранный домен и линкер, который взаимодействует с внеклеточной петлей (ECL2) 7TM
Было также показано, что связывание рецептором агонистов связано с закрытой/активной конформацией VFT участка B1 субъединицы,
а взаимодействие с антагонистами - с открытой/неактивной. Формирование гетеродимера VFT происходит за счет нековалентных
взаимодействий между этими доменами двух субъединиц.
Для трансмембранных доменов также показаны взаимодействия между субъединицами. В неактивном сотоянии наблюдаются
взаимодействия между спралями TM3 (GABAB1) – TM5 (GABAB2) и TM5 (GABAB1) – TM3 (GABAB2). При активации рецептора
происходит ротация доменов, из-за чего начинают взаимодействовать уже спирали TM6 каждой субъединицы.
Как уже было сказано выше сайт связывания агонистов и антагонистов находится в субъединице B1. Этот сайт
образуется аминокислотами из двух долей субъединицы B1 (называются они LB1 и LB2). Аминокислоты, которые
располагаются в LB1 участвуют в связывании как антагонистов, так и агонистов, при этом взаимодействия с остатками
для них обоих очень схожи. 2 аминокислоты, образующие сайт связывания (Trp, Tyr) находятся в LB2. При этом для них выделяют
различные взаимодествия остатков с агонистами и антагонистами.
Остальные особенности устройства рецептора в большей мере связаны с B2 субъединицей, поэтому ее функции пока не будем
затрагивать, если, конечно, при выполнении задания эти знания не понадобятся.
Таким образом из литературы можно сделать следующие выводы о влиянии мутаций на этот рецептор.
При замене аминокислоты в структуре рецептора может произойти:
Может нарушиться целостность как самой субъединицы, так и гетеродимера.
Может нарушиться функция связывания субстрата (здесь мне немного повезло, т.к. выше был упомянут тот факт,
что антагонисты и агонисты связываются одинаковым набором аминокислот, поэтому можно будет рассмотреть
хотя бы одну структуру с любым лигандом).
Может нарушиться локализация или стабильность рецептора при нахождении в липидной оболочке (при
появлении в 7TM полярных аминокислот, направленных в толщу фосфолипидного бислоя).
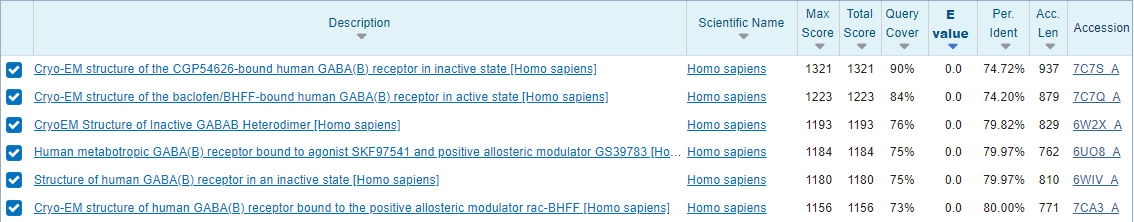
После сбора информации о самом рецепторе был запущен белковый бласт против PDB.
Всего нашлось 28 записей. Все они относятся либо к человеку, либо к дрозофиле. 6 лучших находок (рис. 2.) имеют процент идентичности
больше 74% (наибольшее значение 80%) при этом покрытие больше 73% (наибольшее значение 90%). В принципе это не очень
удивительно, ведь человек и щука не самые близкие родственники. Остальные находки имели либо большие значения покрытия
и маленькие значения идентичности (их мне сложно объяснить), а остальные имели маленькие значения покрытия и различные
значения идентичности от 28% до 80% (это связано по-видимому с тем, что есть структуры лишь отдельных доменов субъединицы
из-за чего и появлялись находки с маленьким покрытием, но хорошим значением идентичности).
Рис. 2. Выдача blastp с параметрами по умолчанию против PDB DB.
На первых парах я решил остановиться на структуре 7C7S из организма Homo sapiens. Выбрал я ее из-за 90% покрытия (лучшее из всех), хоть и с 75%
идентичностью, что не удивительно (см. выше). Более того эта структура содержит в себе обе субъединицы, что выжно для
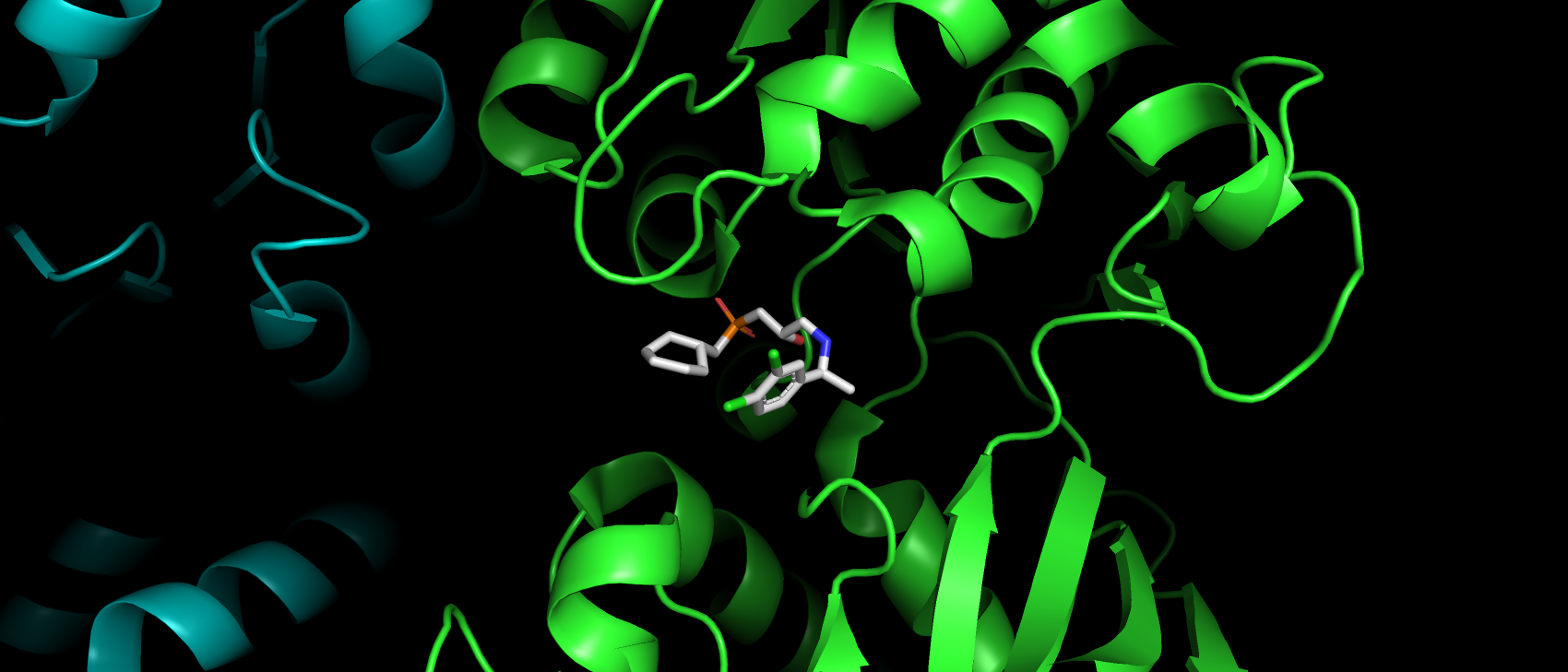
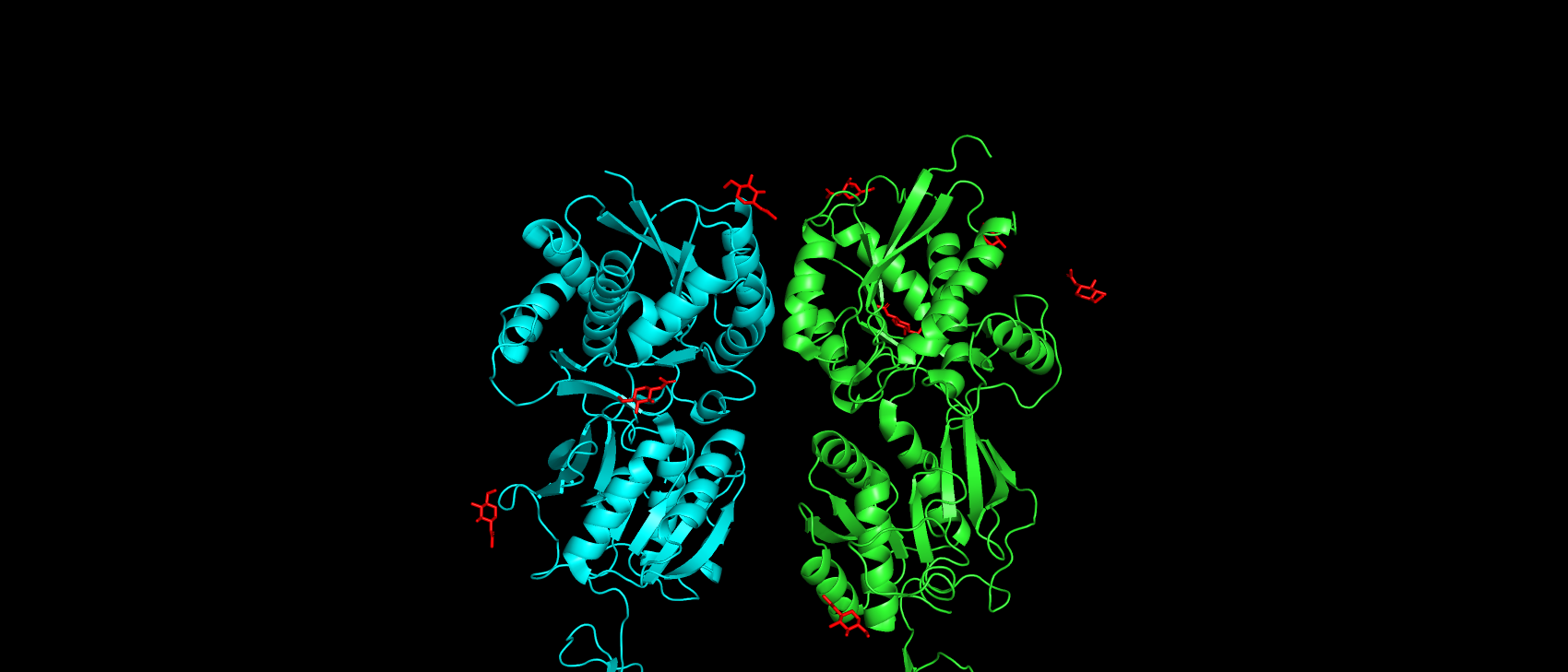
рассмотрения случаев замены аминокислот в участках их взаимодействия, и субстрат-антагонист CGP54626 (рис. 3). Глюкопираноза, которая
также указана в списке лигандов по-видимому не является субстратом рецептора (т.к. облепляют структуру рецептора снаружи и не находится в кармане связывания) (рис. 4).
Стоит отметить, что субстрат имеет полную структуру.
Рис. 3. Субстрат-антагонист CGP54626.
Рис. 4. Расположение глюкопираноз в структуре рецептора.
Нумерация в 7C7S отличается от нумерации в Uniprot на +4.
Первая мутация
Первая мутация: W392H, которая в структуре pdb будет иметь номер 392 + 4 - 1 (-1 из-за одного гэппа, появившегося
в последовательности PDB после выравнивания с последовательностью из Uniprot) = 395.
Обе аминокислоты имеют ароматичные кольца. Оба радикала способны образовывать водородные связи.
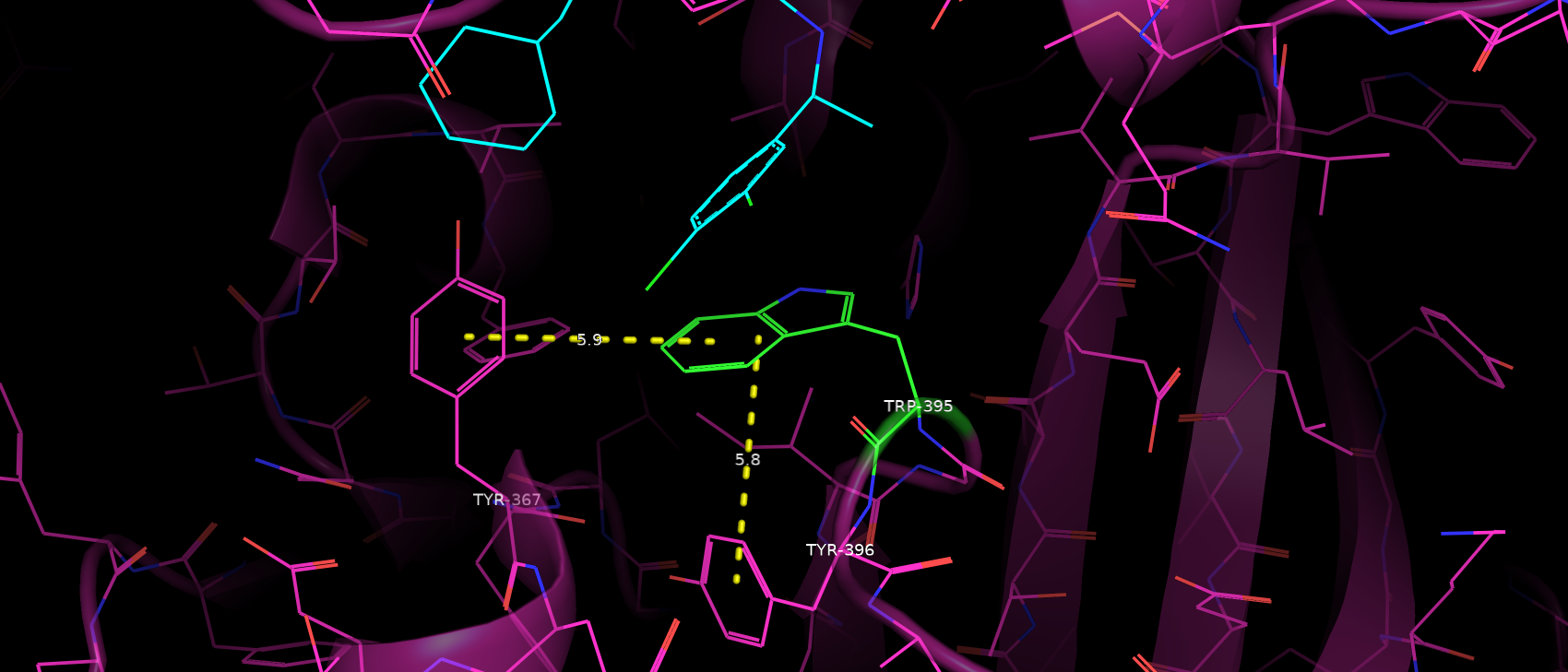
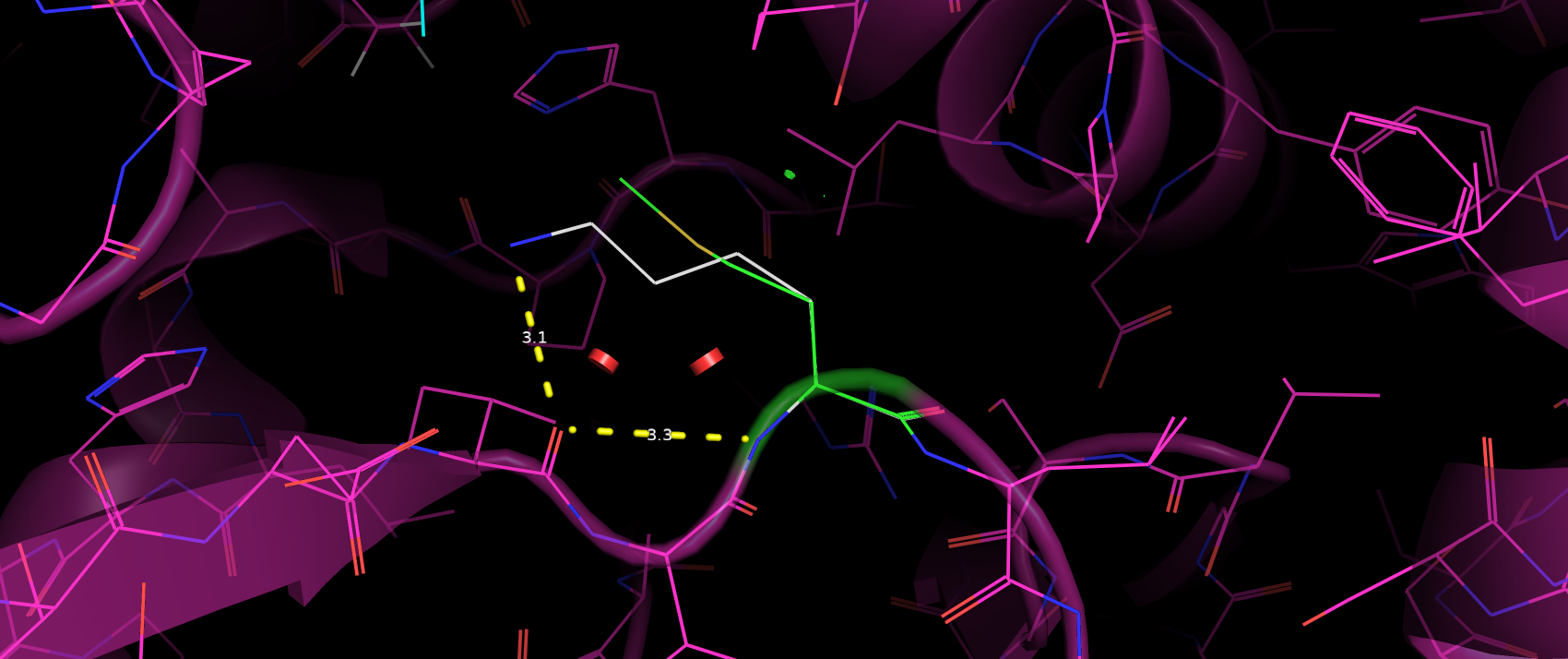
Триптофан 395 не входит с окружающими его аминокислотами ни в какие взаимодействия. Сначала у меня было предположение,
что Trp395 образует пи-стэкинг с Tyr396, однако при расстояние между их кольцами слишком велико (да и по геометрии, кажется,
что тирозин не достаточно повернут в сторону триптофана). С Tyr367 аналогичная ситуация. Таким образом можно сделать вывод,
что Trp395 не участвует в поддержании третичной структуры и фолда в целом (рис. 5).
Рис. 5. Расположение Tyr396 и Tyr367 относительно Trp395.
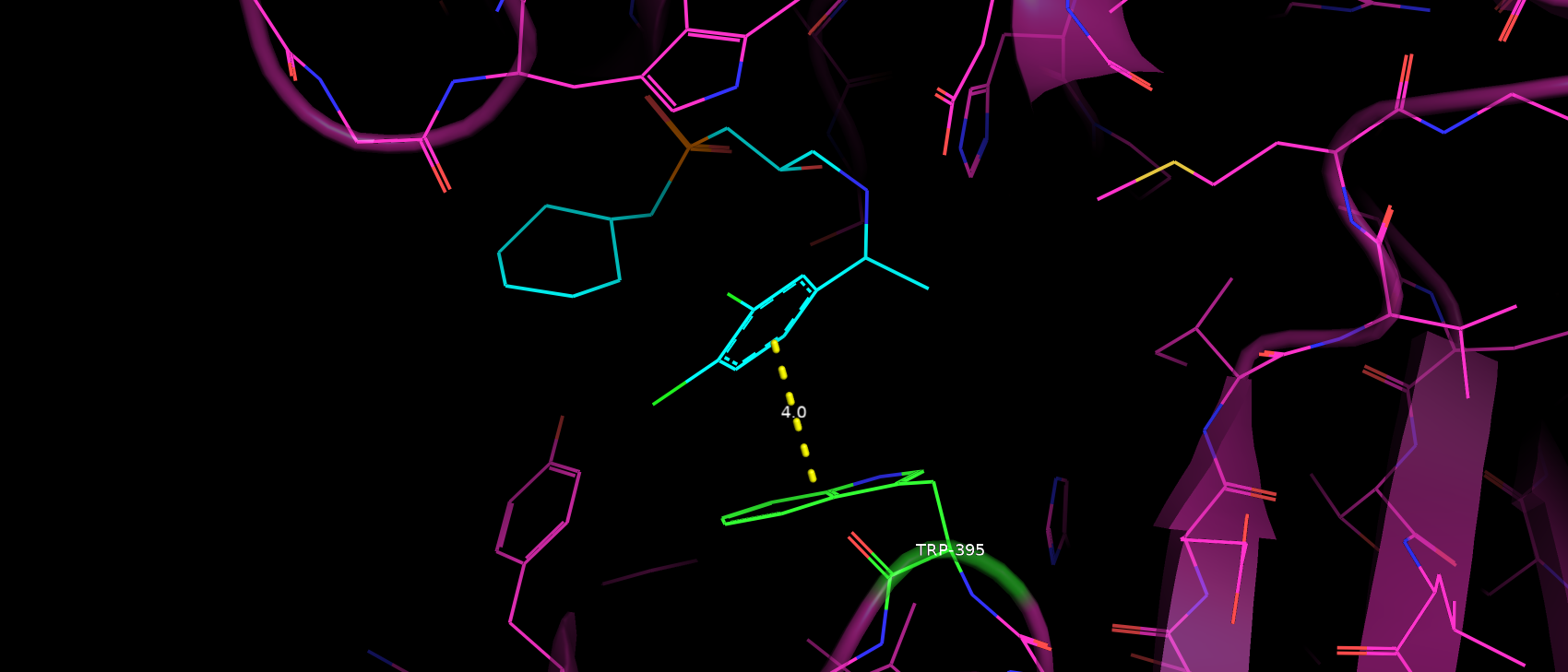
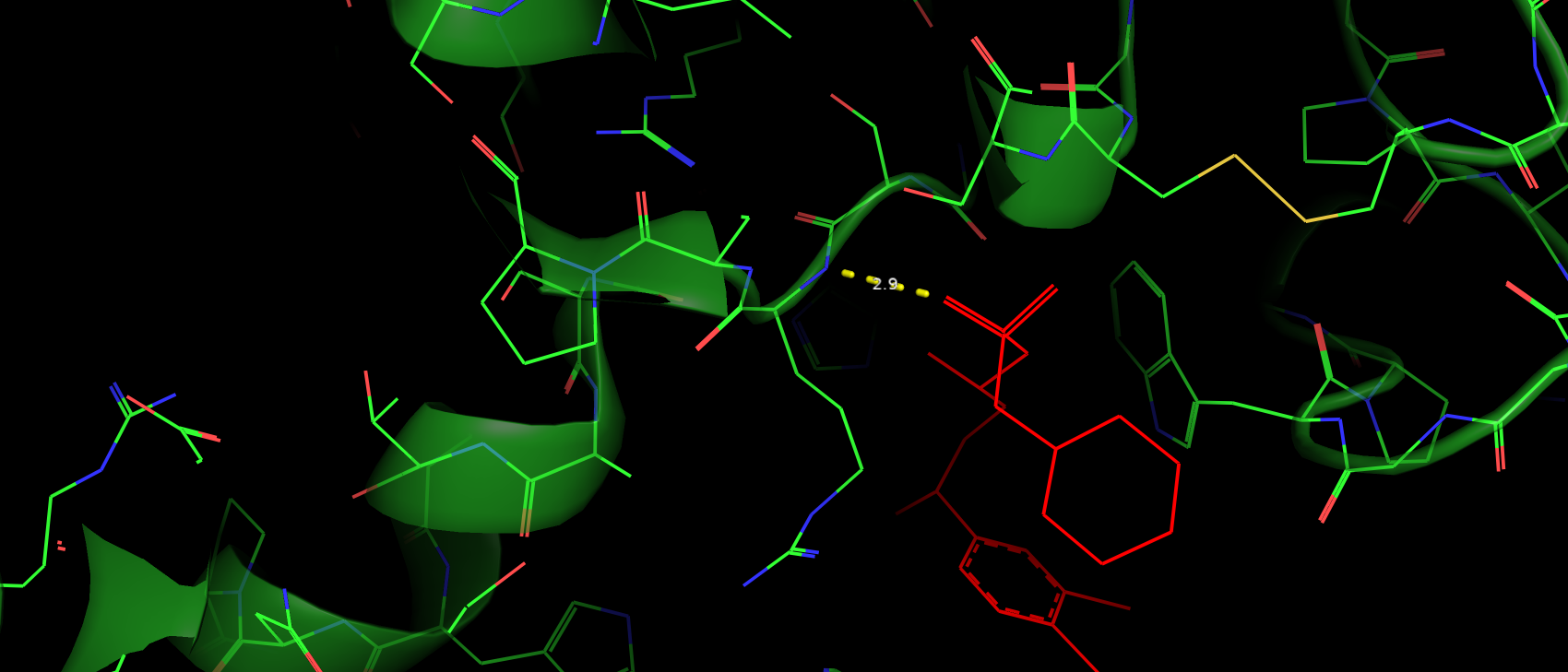
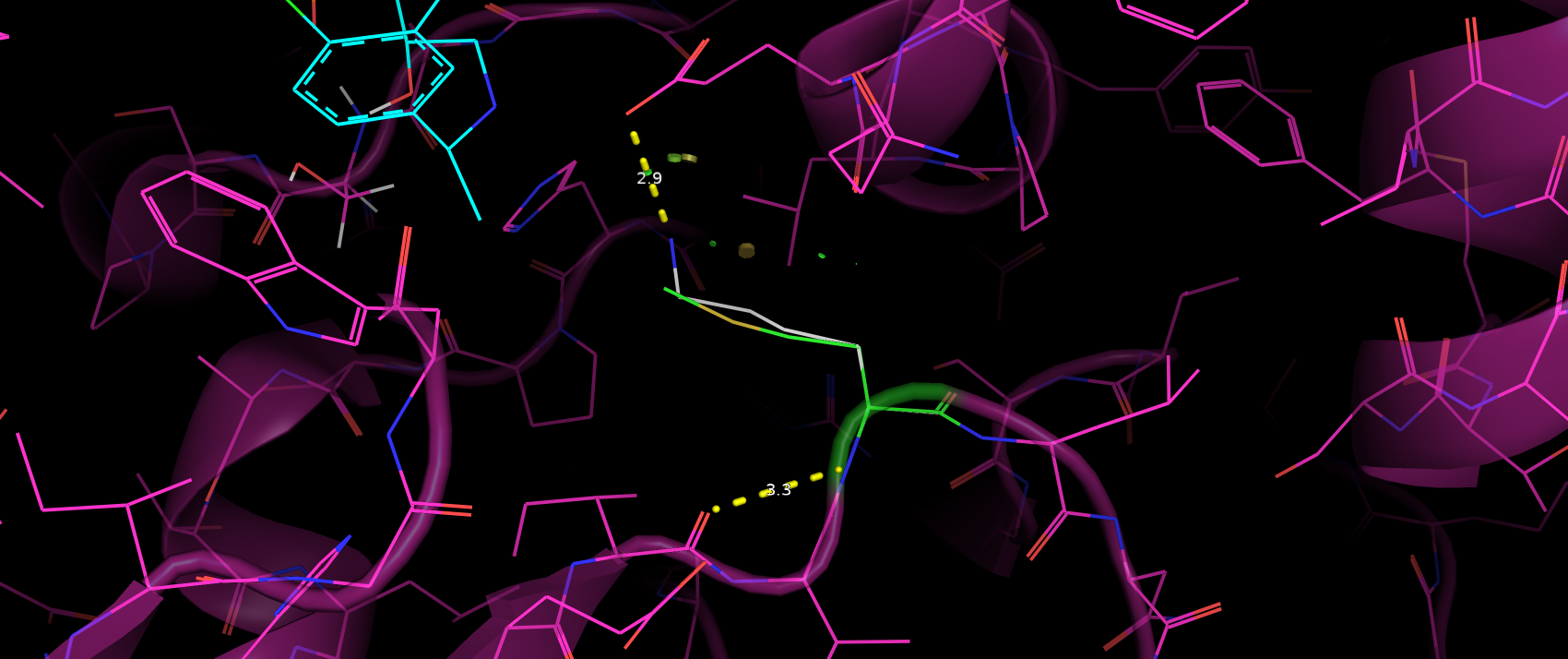
Trp395 скорее всего вступает в стэкинг взаимодействие с ароматичным кольцом субстрата (рис. 6).
Расстояние и геометрия в пределах допустимого, хотя и не идеальные. Таким образом можно предположить, что такое взаимодейстие
не вносит глобального влияния на связывание субстрата.
Рис. 6. Расположение ароматичного кольца субстрата относительно Trp395.
Проведен мутагенез. При этом выбран ротамер с наименьшим стрэйном = 12,55.
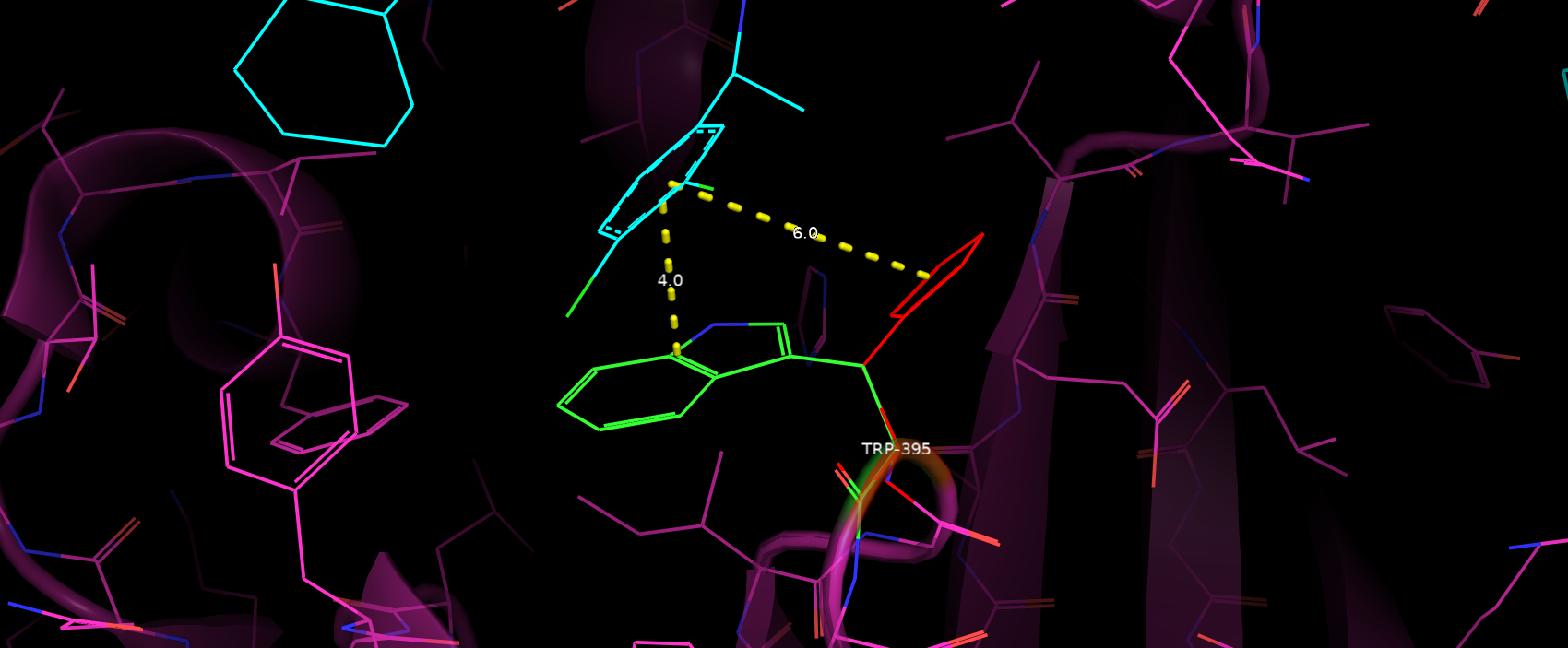
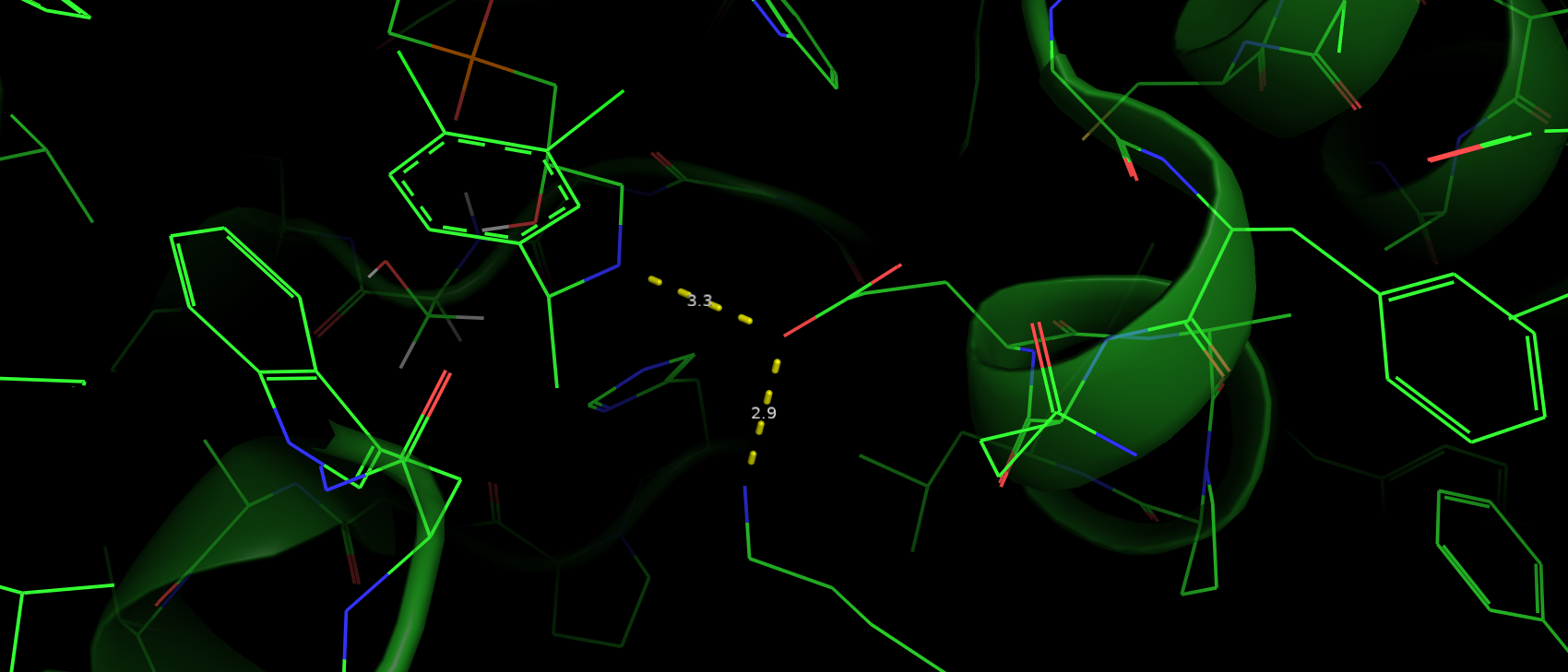
Мутантаный гистидин также не образует новых водородных связей с окружением. Помимо этого из-за сильного отдаления
от ароматичного кольца субстрата теряется стэкинг взаимодействие (рис. 7).
Рис. 7. Расположение ароматичного кольца субстрата относительно мутантного His (красный цвет).
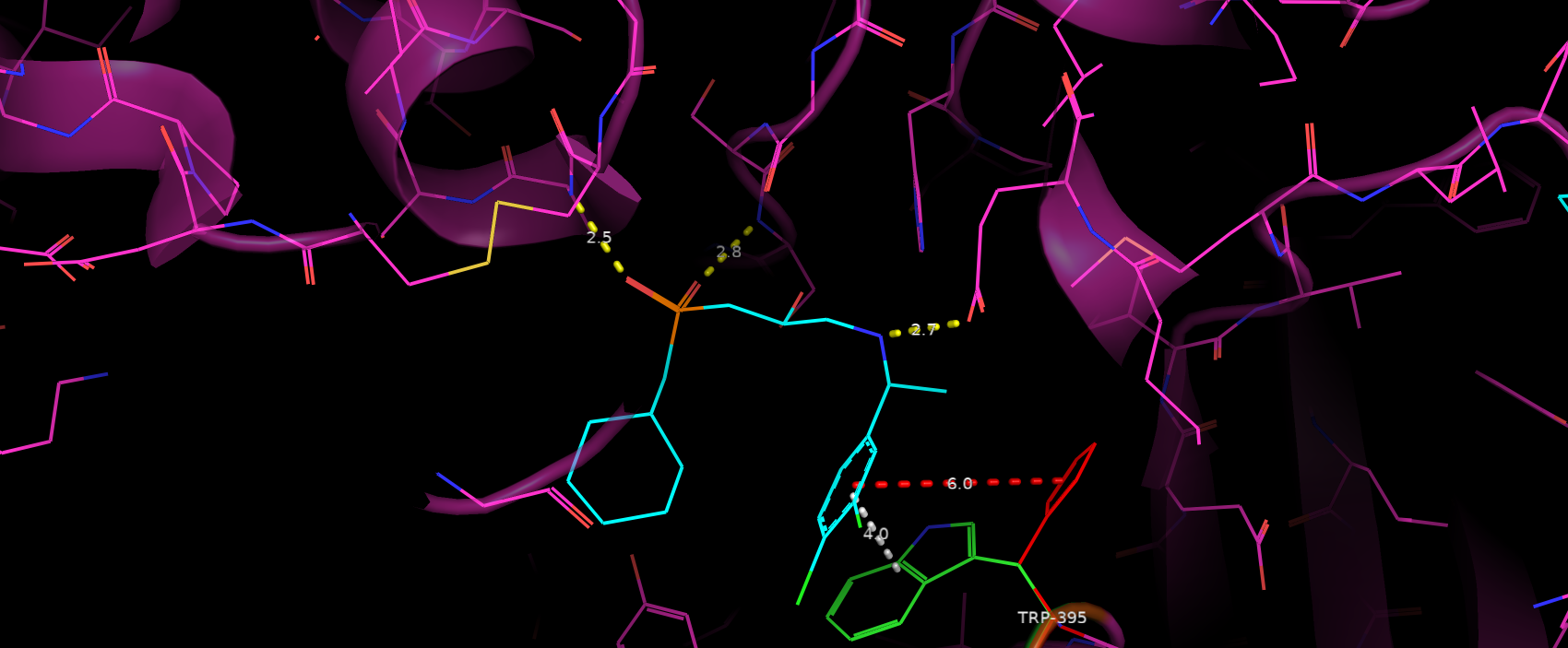
Так как фолд самой структуры при такой замене не изменился, значит другие взаимодействие рецептора с субстратом
не изменились (рис. 8). Сохранились 3 водородные связи, которые скорее всего несут основную роль в связывании субстрата.
Таким образом я бы сделал вывод, что данная мутация имеет слабо негативный эффект на связывание субстрата.
Рис. 8. Взаимодействия аминокислот сайта связывания с субстратом: желтый - водородные связи,
белый - стэкинг взаимодействие до мутации, красный - утерянное взаимодействие после мутации.
Вторая мутация
Вторая мутация: S267R. В структуре pdb серин будет иметь номер 267 + 4 - 1 (аналогично первой мутации) = 270.
Обе аминокислот имеют гидрофильный радикал. Способны к образованию водородных связей. Аргинин положительно заряжен и
потому может участвовать в образовании солевых мостиков. Аргинин имеет куда более длинный радикал по сравнению с серином.
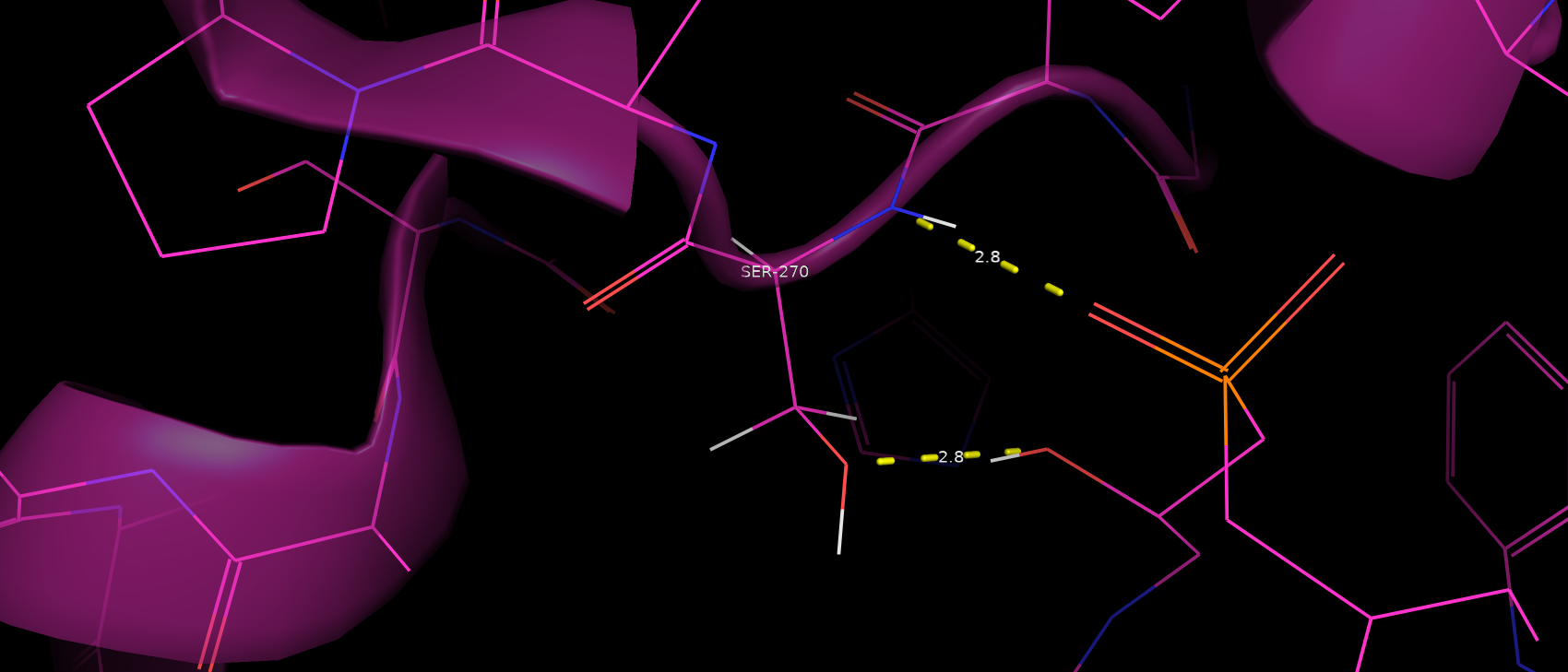
Серин 270 оказался довольно интересным для рассмотрения. При рассмотрении первой мутации я забыл упомянуть про водородную
связь не только между остовом серина 270, но и его радикала с субстратом (расстояние здесь хорошее, а геометрия на грани,
хотя для серина мы отмечали возможность образования водородных связей даже при угле 90 градусов) (рис. 9).
Рис. 9. Водородные связи между серином 270 и субстратом.
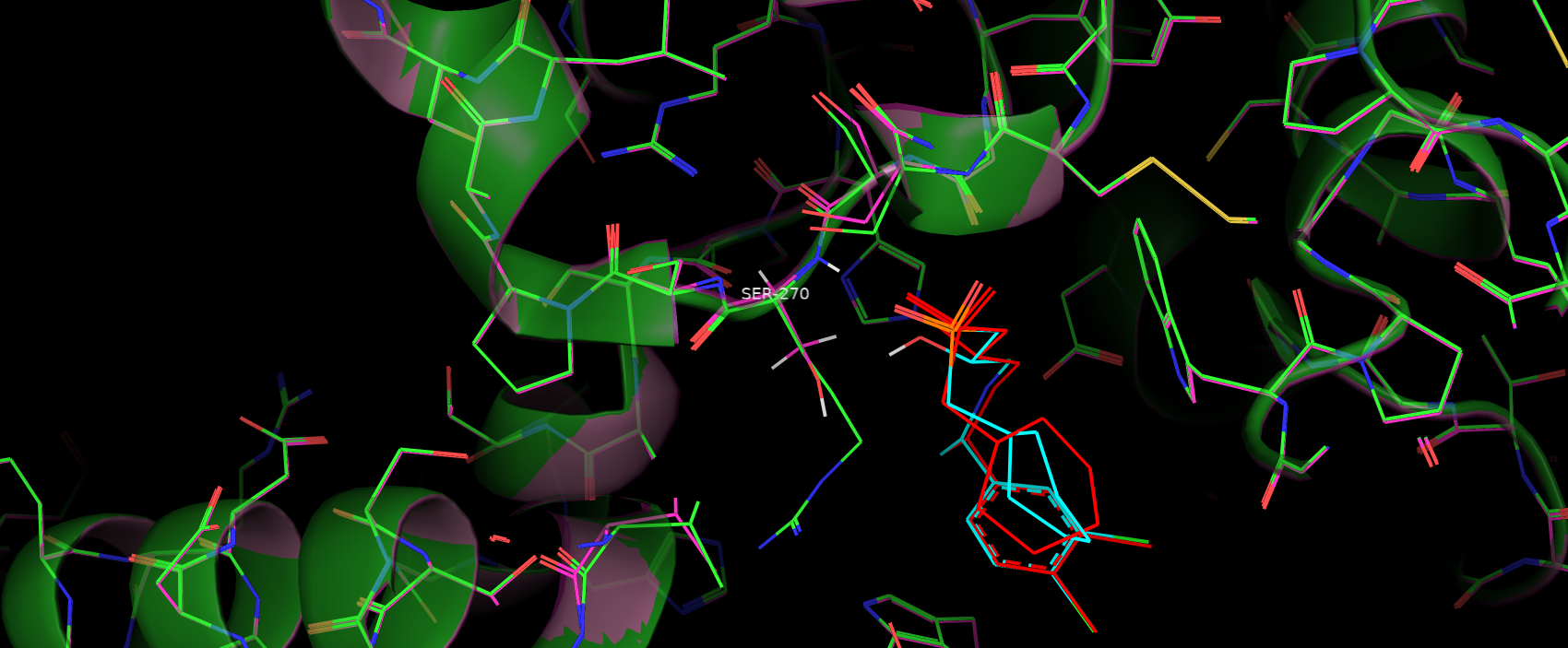
При замене серина на аргинин (после мутации выбран ротамер с самым маленьким стрэйном = 21.86 и проведен скалптинг)
структура почти не изменилась, как и положение субстрата в кармане связывания (рис. 10)(произошел только поворот вокруг 1-ой связи
из-за чего развернулось неароматичное кольцо субстрата). Поэтому осталась почти без изменений водородная связь между
остовом Arg 270 (рис. 11).
Рис. 10. Наложение структур до и после мутации, видно, что остов не поменял своего положения
из чего можно заключить, что другие взаимодействия, перечисленные на рис. 8, сохранились в том числе с оставом
270 аминокислоты. Видно, что
не ароматичное кольцо субстрата поменяло свое положение. Красный цвет - положение субстрата после мутации
Рис. 11. Водородная связь между остовом аргинина 270 и субстратом.
Однако теперь утрачена водородная связь с боковым радикалом, при этом новых взаимодействий боковой радикал
Arg не образует.
Таким образом получается, что такая мутация не вносит изменений в третичную структуру рецептора и его фолд,
основное количество взаимодействий (3 из 4 водородных и стэкинг) сохраняются. Получается, что подобная замена
может привести к увеличению энергии связывания субстрата, что может привести к уменьшению специфичности, афинности рецептора.
Я бы сказал, что данная мутация несет негативный эффект.
Третья мутация
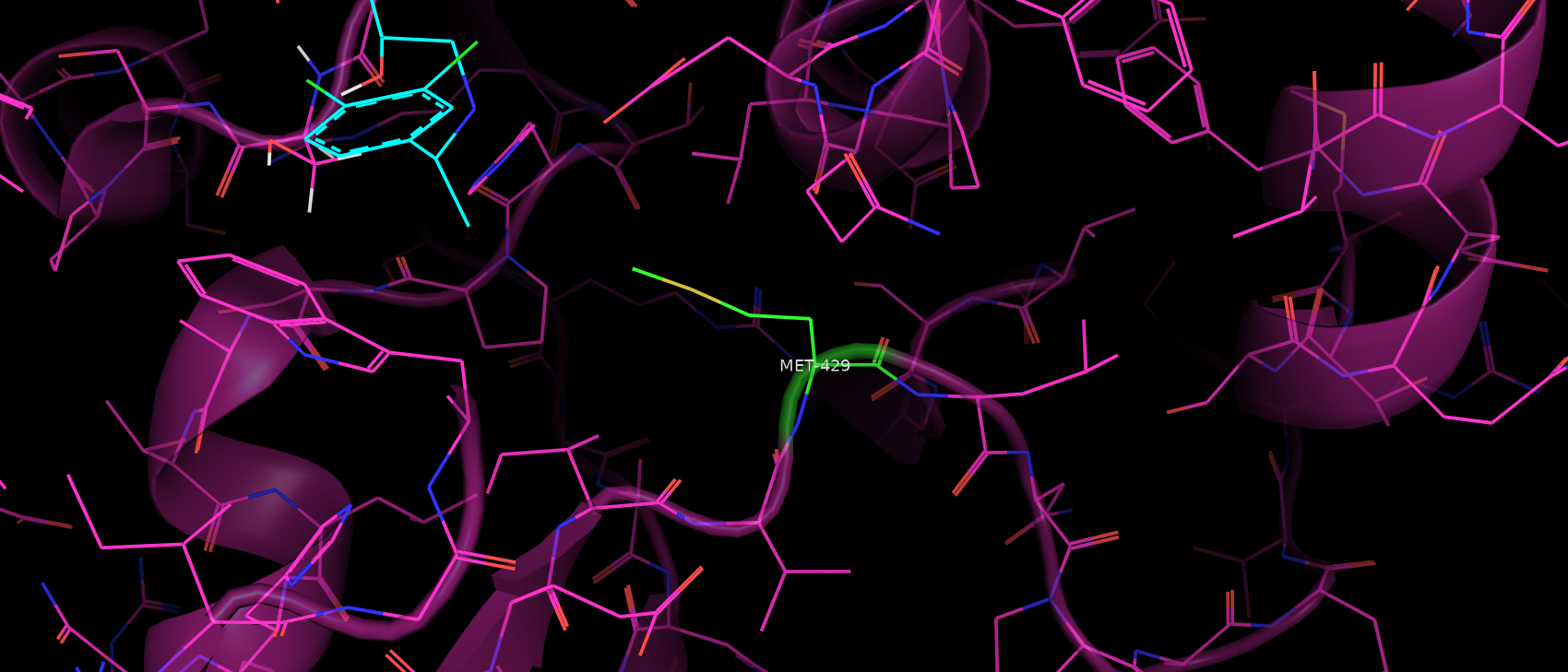
Третья мутация: M426K. Метионин в структуре PDB будет иметь номер 426 + 4 - 1 = 429.
Аминокислоты разные по химическим свойствам. Лизин может образовывать водородные связи, его радикал гидрофильный.
Метионин в данном случае скорее всего выполняет роль заполнения пространства. При этом вокруг него довольно много
гидрофобных аминокисотных радикалов, с которыми он вероятно имеет гидрофобные взаимодействия. Возможно эти взаимодействия
могут влиять и на связывание субстрата.
Рис. 12. Расположение метионина 429 относительно субстрата.
При проведении мутации из 20 ротамеров самые маленькие значения стрэйна были у 2 (рис. 13, 14).
Рис. 13. Ротамер 1 лизина со стрэйном = 18.76
Рис. 14. Ротамер 2 лизина со стрэйном 19.14.
Оба ротамера имеют маленький стрэйн, однако, ротамер 1 не внесет никакого изменения ни в структуру, ни в связывание
субстрата, так как не образует никаких новых взаимодействий.
Ротамер 2 выглядит куда интереснее, так как он может образовать водородную связь с Glu466, который в свою очередь уже
взаимодействует с субстратом.
Однако после проведения скалптинга, получилось, что Glu466 отдалилась от субстрата (рис. 15).
Однако, если принять, что водородные связи могут иметь рассотяние до 3.5 Å, то в таком случае водородная связь останется.
Рис. 15. Положение Glu466 до (фиолетовый) и после (зеленый) мутации.
Рис. 16. Водородные связи Glu466 после мутации.
Таким образом ответ здесь может отличаться от того, какой же все-таки ротомер действительно был бы в структуре после
мутации. Но в любом случае такая мутация либо не имеет никакого влияния (в обоих случаех), либо имеет очень слабый негативный
эффект при ротамере 2.