
В рамках задания мною был выбран µ-кристаллин сумчатых. Этот белок описан в данной статье.
Для поиска дальних родственников был выбран белок Кетиминредуктаза мю‑кристаллин (Ketimine reductase mu-crystallin). Идентификатор в Swiss‑Prot: Q28488 (CRYM_MACFL). Функция: катализирует восстановление иминосвязей (кетиминредуктазная активность), связывает NADPH. Длина последователньости составляет 314 аминокислот.
Вероятный предковый белок - аспартокиназа II / гомосериндегидрогеназа II. Это бифункциональный фермент, играющий ключевую роль в аспартатном пути биосинтеза аминокислот. Имеет три домена: N-концевой домен: катализирует первую реакцию пути — фосфорилирование L-аспартата до 4-фосфо-L-аспартата, используя АТФ, центральный линкерный домен: соединяет два каталитических домена и C-концевой домен, который катализирует третью реакцию пути — НАДФ-зависимое восстановление L-аспартат-4-полуальдегида до L-гомосерина. Этот фермент относится к надсемейству Орнитин-циклодеаминаза/µ-кристаллин. И в статье было указано, что µ-кристаллин гомологичен не всему этому огромному белковую комплексу, а конкретно C-концевому домену гомосериндегидрогеназы.
С помощью PSI-BLAST в базе данных Swiss-Prot проводился поиск родственных последовательностей. Итого было проведено 5 итераций, после проведение дополнительных итераций было остановлено, так как количество новых последовательностей продолжало увеличиваться, а большей конкретики находки не давали, лишь подтверждали предыдущие.
По итогам первой итерации всего нашлось 36 последовательностей. Среди них первые пять значимых находок с E-value = 0 имеют высокую идентичность, одинаковую длину (313‑314 а.о.) и выполняют функцию кетиминредуктазы / НАДФ‑зависимого связывания тиреоидного гормона. Остальные (31) находка являются отдаленными гомологами и среди них все являются членами того же надсемейства ферментов метаболизма аминокислот.
Во второй итерации количество находок увеличилось до 456. Среди находок присутствуют ферменты, функционально близкие к предковому (аланиндегидрогеназа, иминосукцинатредуктаза, орнитин‑циклодеаминазы), а также множество глутамил‑тРНК‑редуктаз, демонстрирующих структурное сходство с µ‑кристаллином, имеющий тот самый НАДФ‑связывающий домен.
В третьей итерации число находок вновь возросло, уже до 1041 последовательности. Огромное количество глутамил‑тРНК‑редуктаз (GluTR) – сотни гомологов из протеобактерий, фирмикут, цианобактерий, архей и даже эукариот (растения, грибы). Эти белки структурно являются НАДФ‑зависимыми оксидоредуктазами.
Число находок - 1593. В четвёртой итерации помимо множества GluTR появились: шикиматдегидрогеназы (SDH) , которые катализируют НАДФ-зависимое окисление шикимата и является ключевым ферментом биосинтеза ароматических аминокислот. Их появление указывает на сохранение НАДФ-связывающего домена, общего с µ‑кристаллином. И еще аденилгомоцистеиназы (AHCY) и их гомологи. Эти фермент также содержат НАДФ-связывающий домен и демонстрируют структурное сходство с µ‑кристаллином.
Количество находок в пятой итерации возросло до 2387 последовательностей. Количество уникальных семейств белков практически не увеличилось. Вместо этого, произошло значительное расширение уже найденных семейств за счет большого числа гомологов из различных таксономических групп (бактерии, археи, эукариоты).
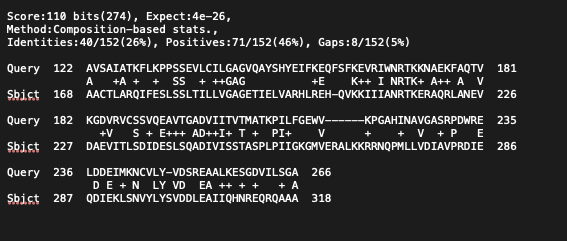
Проведенный анализ подтверждает гипотезу о происхождении µ-кристаллинов сумчатых от ферментов метаболизма аминокислот. В ходе поиска PSI-BLAST была выявлена эволюционная связь между белком хрусталика и обширным суперсемейством НАДФ-зависимых оксидоредуктаз (семейства орнитин-циклодеаминазы/гомосериндегидрогеназы). Появление в поисковой выдаче таких ферментов, как аланиндегидрогеназы, глутамил-тРНК-редуктазы и шикиматдегидрогеназы, говорит нам о высокой консервативности каталитического домена.