Базы данных KEGG и GO
Мой список ID:
- PLA2G12A
- PLA2G12B
- PLA2G2E
- PLA2G2F
- PLA2G2C
- PLA2G4E
- PLA2G2D
- PLA2G4F
- PLA2G2A
- PLA2G4C
- PLA2G4D
- PLA2G4A
- PLAAT3
- ACOX3
- ACOX1
- PLA2G10
- PLA2G1B
- PLA2G3
- PLA2G5
- PLA2G6
- PLB1
- ACAA1
- FADS2
Он содержит 25 ID, но частью баз данных JMJD7-PLA2G4B не распознается у человека. JMJD7 и PLA2G4B гены кодируют разные белки, но находятся в одном локусе. JMJD7 функционально имеет мало отношения к остальным белкам списка.
Исследуем функции белков, кодируемых этими генами, как метаболические, так и молекулярные, попытаемся определить общую для всех нишу обмена веществ. Все данные по Homo sapiens. Использованы сервисы GO enrichment analysis и Reacfoam, позволяющие упорядочить и удобно просмотреть и сопоставить аннотирование генов по функциям и другим свойствам их белков (в нашем случае). Оба сервиса позволяют упорядочить полученные результаты, опираясь на обогащение списка генов определенными тегами, благодаря чему легко и надежно определяются пересекающиеся и входящие друг в друга множества тегов. Reacfoam кроме этого предоставляет несколько визуальных инструментов, ускоряющих исследование функций белков и позволяющих их кратко представить.
GO enrichment analysis
Полученные результаты отсортированы по возрастанию FDR, чтобы обнаружить наиболее значимые теги, после чего вручную были определены самые значимые функциональные подмножества: например транспорт арахидоновой кислоты является подмножеством транспорта органических кислот, имея при этом столько же вхождений из списка.
Исходя из полученных результатов можно сказать, что список генов полностью относится к липидному обмену, причем 22 из 24 искомых генов относятся к катаболизму. К обмену арахидоновой кислоты относятся 13 генов. Другое значимое подмножество (20 вхождений) относится к обмену фосфолипидов: фосфатидилхолин, фосфатидилинозитол.
В группу обмена арахидоновой кислоты вошли только PLA2G… гены (Phospholipase A2 G), кроме:
- PLA2G4E
- PLA2G4C
- PLA2G4D
- PLA2G4B
- PLA2G6
Которые относятся только к обмену фосфолипидов.
В группу катаболизма липидов не вошли только гены PLA2G3 (секреторная фосфолипаза) и FADS2(Acyl-CoA 6-desaturase).
По молекулярной функции все белки обладают каталитической активностью, 20 из 24 оказались A2 фосфолипазами (даже PLB1). У шести PLA2 белков отмечается еще и PLA1 активность. Несмотря на то, что все белки отнесены к обмену липидов и имеют каталитическую активность, только 17 из них по тегам молекулярной функции связываются с липидами непосредственно, возможно, некоторые субстраты имеют высокую полярность (вероятно, ацил-CoA субстраты и незамещенные по 1 атому глицерина фосфолипиды).
По (выделяющейся) локализации 4 белка относятся к микротельцам (пероксисомам), расположение остальных вполне обычно и имеет или высокое значение P-value или близкое к единице обогащение списка.
Reacfoam
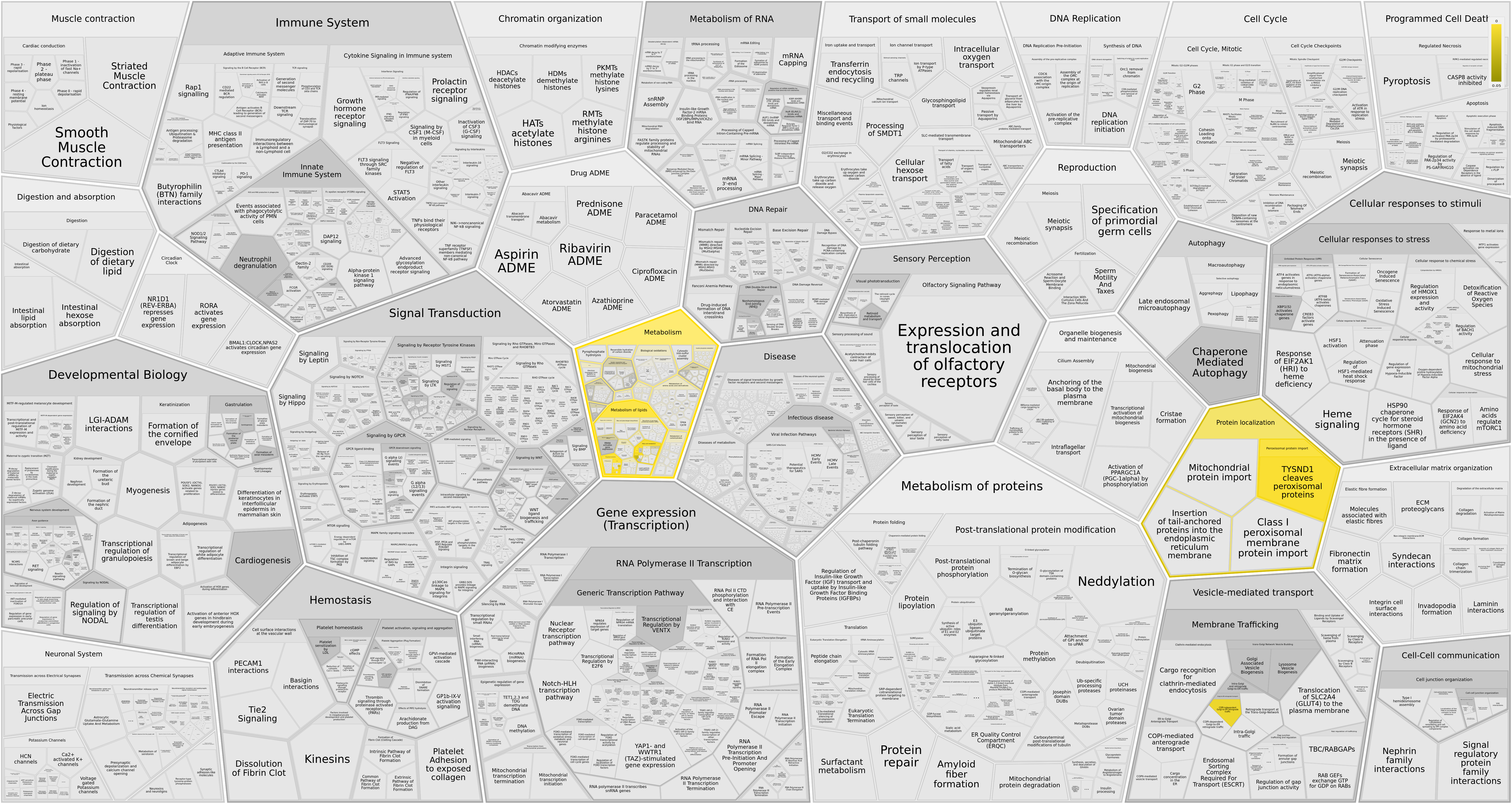
Основные категории, к которым относились указанные гены:
- Acyl chain remodeling PG, PI, PS, PE, PC (фосфатидилглицерола, фосфатидилинозитола, фосфатидилсерина и т.д.)
- Synthesis of PA
- Phospholipid metabolism (19 вхождений)
- Glycerophospholipid biosynthesis (19 вхождений)
Новая информация получается из категорий:
- Hydrolysis of lysophosphatidic acid (6)
- Alpha-linolenic and linoleic acid metabolism (3, из них только одна десатураза)
Что расширяет спектр липидов, в метаболизме которых участвуют белки из списка.
Reactfoam подтверждает данные GO о локализации в пероксисомах трех из четырех белков: ACOX1, ACAA1, ACOX3, относя их к импортируемым в пероксисому белкам. Эти белки по представленной карте метаболических процессов являются участниками бета-окисления жирных кислот.
Вывод
По результатам инструментов анализа обогащения можно сразу заметить, что белки с A2 фосфолипазной активностью имеют каждый пересекающееся с другими такими белками множество субстратов, задействованных в различных процессах, преимущественно завязанных на ремоделирование мембранных липидов и освобождение из триглицеридов субстратов для синтеза эйкозаноидов. Из пересечения их субстратов следует сложное, нелинейное описание их взаимосвязанной регуляции и эволюции.
Белки без фосфолипазной активности ACOX1, ACAA1, ACOX3 являются участниками бета-окисления жирных кислот, но, вероятно, могут выполнять и другие функции, в зависимости от локализации в клетке (пероксисома, митохондриальный матрикс), в том числе приводя к образованию ненасыщенных липидов различного назначения. Кроме того, ненасыщенные жирные кислоты, по некоторым данным, легче вступают на путь бета-окисления (хотя и нуждаются в насыщении двойной связи на определенном этапе). FADS2 же является непосредственно десатуразой и участвует в модифицировании полиненасыщенных жирных кислот.
Таким образом можно выделить наиболее общую, но не единственную, роль всех без исключения генов списка: обмен полиненасыщенных жирных кислот.
Кроме того, можно заметить, что у Reacfoam, вероятно, более качественное аннотирование, которое позволяет не потерять информацию о множестве различных субстратов фосфолипаз A2, не сводя все к высвобождению арахидоновой кислоты.