Алгоритмы множественного выравнивания.Pfam
Различия во множественных выравниваниях, построенных разными программами
Для выполнения данного задания я взяла пять гомологичных белков (из практикума 10): A9A135, C1F2D2, P9WMJ8, Q85FW4 и A0T0H7. На рис.1 можно увидеть выравнивания этих белков, выполненные программами Mafft, Muscle и Tcoffee (на рисунке в том же порядке).Теперь рассмотрим различия между выравниваниями. Вначале выранивания одинаковы, отличия начинаются, когда появляется много гэпов.
Рассмотрим позицию 93. В первым выравнивании под Tyr93 (во второй последовательности) находится Trp93(в третьей последовательности), в двух других выраниваниях под Tyr93 находятся гэпы, а Trp сдвинут на 81 позицию.
В первом выранивании над Val95 (вторая последовательность) находится Phe95 (первая последовальность), во втором и третьем выравниваниях над Val95 гэп, а Phe95 расположен над Val107.
В позиции 293 в первых двух выравниваниях под Ser293 (первая последовательность) находится Thr293 (вторая последовательность), а в последнем выранивании Thr293 находится под под Ser294. Кстати, из-за добавления гэпов в 293 и 295 позициях третье выравнивание сдвигается после этих позиций на одну позицию относительно первых двух.
Таким образом, мы видим, что выравнивания, построенные разными программами, одинаковы в тех местах, где находятся абсолютно/функционально консервативные или высоко консервативные колонки, и могут отличаться там, где необходимо использовать гэпы (в этих местах в последовательностях произошли делеции/вставки).
Рис.1. Множественное выравнивание белков, построенное разными программами

Различные доменные архитектуры, содержащие один и тот же домен
На рис.2 приведена доменная архитектура белка белка 3-оксоакцил-[АПБ]-синтаза (BAC70085.1). Белок имеет два домена: ACP_syn_III (обозначен зеленым, в моем белке неполный, занимает 106 - 182 а. о.) и ACP_syn_III С (обозначен красным, 239 - 330 а. о.). ACP_syn_III участвует в катализе определенных реакций и присущ ферментам, относящихся к семейству трансфераз. Beta-Ketoacyl-acyl-carrier-protein synthase III (ACP_syn_III) участвует в диссоцииации систем биосинтеза жирных кислот, которая происходит в растениях и бактериях. [1] Функция второго домена ( ACP_syn_III С) не ясна, однако можно предположить, что он помогает первому домену, внося свой вклад в катализ.Рис.2. Архитектура белка 3-оксоакцил-[АПБ]-синтаза

Для поиска различных архитектур я выбрала домен ACP_syn_III своего белка. Затем я провела поиск в Pfam по этому домену и нашла много разных архитектур с ним. Далее я расскажу о трех из них.
1. Белок A0A0J7KII5_9BACT [Chitinispirillum alkaliphilum] помимо тех же доменов, что и мой белок, имеет еще Biotin_lipoyl (обозначен бардовым). Этот биотин-липоль домен имеет консервативный остаток лизина, который связывает биотин или липоевую кислоту. Биотин играет роль катализатора в некоторых реакциях переноса карбоксильных групп и с помощью амидной связи ковалентно присоединен к остатку лизина в ферментах, для которых он или липоевая кислота играет роль кофермента. [2] Доменная архитектура белка представлена на рис.3, последовательность доменов следующая: Biotin_lipoyl, ACP_syn_III, ACP_syn_III_C.
С такой доменной архитектурой нашелся лишь один белок.
Рис.3. Архитектура белка A0A0J7KII5_9BACT

2. У белка A0A0L0KAM1_9ACTN [Streptomyces acidiscabies] тоже присутствуют те же домены, что и в 3-оксоакцил-[АПБ]-синтазе, но есть еще и PP-связывающий домен (обозначен желтым). Он участвует в катализации реакции переноса электронов от NADH к коферменту Q10 (CoQ10). [3] Доменная архитектура белка (рис.4): ACP_syn_III, ACP_syn_III_C, PP-binding.
С такой доменной архитектурой нашлось 10 последовательностей.
Рис.4. Архитектура белка A0A0L0KAM1_9ACTN
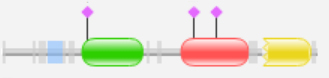
3. А вот в белке R4Z3W8_9ACTN [Candidatus Microthrix parvicella RN1] архитектура (рис.5) такая: ACP_syn_III, Thiolase_C. Тиолаза_С присутствует в таких ферментах, как бета-кетоацил-АТФ синтетаза, которая участвует в синтезе жирных кислот, приводя к образованию ацетоацетилу АТФ. Бета-кетоацил-АТФ синтетаза является весьма консервативным ферментом, который обнаруживается практически у всего живого на земле в качестве домена в синтетазе жирных кислот (FAS). FAS имеет два типа: тип I и II. Тип I находится у животных, грибов и «низших эукариоты». Тип II находится у прокариот, а также в пластидах и митохондриях. [4]
С такой архитектурой было найдено 3 последовательности.
Рис.5. Архитектура белка R4Z3W8_9ACTN
