Проверка дерева цитохромов b
В этом задании было необходимо повторить работу по реконструкции дерева цитохромов b,
описанную в статье
Early Evolution of Cytochrome bc Complexes (Schütz et al., 2000).
Представленное в статье филогенетическое дерево цитохромов (Рис. 1) было реконструировано с методом
neighbour-joining с использованием коррекции Кимуры для многократных замен (Kimura's correction for
multiple substitutions). Гэпы были исключены. В качестве внешней группы были взяты белки архей.
Цифры на узлах соответсвуют частоте их встречаемости (в %) в 5000 бутстрэп-репликах.
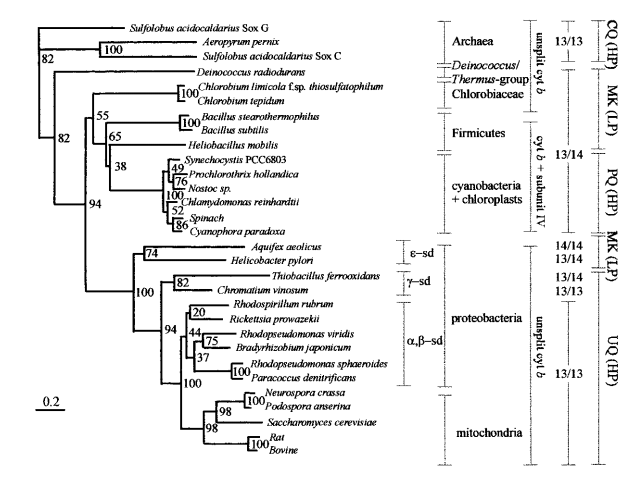 |
| Рис. 1. Филогенетическое дерево цитохромов b (Schütz et al., 2000) |
Используя последовательности белков из указанного файла, я провела собственную реконструкцию филогенетического дерева. Для начала последовательности были открыты в JalView, и программой Muscle было построено их выравнивание (Рис. 2).
 |
| Рис. 2. Выравнивание последовательностей цитохромов b |
Я открыла выравнивание в Mega методом Analyze и построила первое дерево по тому же методу, что и в статье (neighbour-joining c бутстрэп анализом в 5000 реплик и моделью замен p-distance). (Рис. 3)
 |
| Рис. 3. Филогенетическое дерево цитохромов b, Neighbour-Joining, Bootstrap consensus |
Затем было построено еще два дерева с использованием методов Minimal Evolution и Maximum Parsimony. Для каждого из них также был проведен бутстрэп-анализ с 5000 репликами и использована модель замен p-distance.
 |
| Рис. 3. Филогенетическое дерево цитохромов b, Minimal Evolution, Bootstrap consensus |
 |
| Рис. 3. Филогенетическое дерево цитохромов b, UPGMA, Bootstrap consensus |
Во-первых, следует отметить, что построенное таким же методом дерево по топологии хоть и очень схоже, но все же в некоторых местах отличается от представленного в статье. Например, в дереве из статьи есть ветвь, отделяющая Aquifex aeolicus и Helicobacter pylori от остальных, а в моем дереве ее нет. Величины поддержки бутстрэп на совпадающих ветвях также иногда отличаются, хоть и не слишком сильно. Например, ветвь, отделяющая Nostoc sp и Prochlorothrix hollandica в моем дереве имеет поддержку 80, а в статье - 76.
Все три дерева, построенные мной, по топологии совпадают, имеются лишь совсем незначительные различия в величинах бутстрэп-поддержки отдельных ветвей.
На основании построенных деревьев сложно подобрать укоренение, которое было бы однозначно правильным. Одним из возможных можно считать укоренение в ветвь, отделяющую архей от эубактерий (Sulfolobus solfataricus (SoxG), Sulfolobus solfataricus (SoxC), Aeropyrum pernix K1). Именно такое укоренение было предложено в статье. Однако возможны и другие варианты, например в случае, если три архейные последовательности - результат горизонтального переноса от предков эубактерии Deinococcus к предкам архей Sulfolobus и Aerophylum.