ГТФазы семейства Rho служат бинарными молекулярными «переключателями», которые включены или выключены в ответ на различные внеклеточные стимулы. Rho белки в состоянии, связанном с ГТФ (активное состояние), взаимодействуют с разнообразными эффекторами и передают сигнал, который может приводить к перестройкам актинового цитоскелета, регулировать транскрипцию генов, трафик через мембрану и другие функции. После гидролиза молекулы ГТФ Rho белки снова возвращаются в базальное неактивное состояние. Переход Rho белков в одно из двух состояний опосредован работой 3 семейств регуляторных белков, среди которых нас особенно интересует белок, активирующий Rho ГТФазу (RhoGAP).
RhoGAP домен, которым он связывается с ГТФазой, состоит из 9 альфа-спиралей, и на его петле находится высоко консервативный остаток Arg. Считается, что при связывании G белок поворачивается по отношению к RhoGAP, и эта конформационная перестройка помещает каталитический Arg RhoGAP в активный сайт Rho ГТФазы и так стабилизируется переходное состояние при протекании реакции. [1]
Подготовка
Мне был предложен белок с индентификатором A0A6P6KFY1. Это белок, подобный белку 1, активирующему ГТФазу (rho GTPase-activating protein 1-like) из организма Carassius auratus (Золотая рыбка). Конкретно этот белок изучен слабо, поэтому была предпринята попытка найти похожие белки, для которых показана структура. Для этого последовательность аминокислотных остатков (длина 434 а.о.) этого белка была заружена в Blast и проведен поиск по PDB.
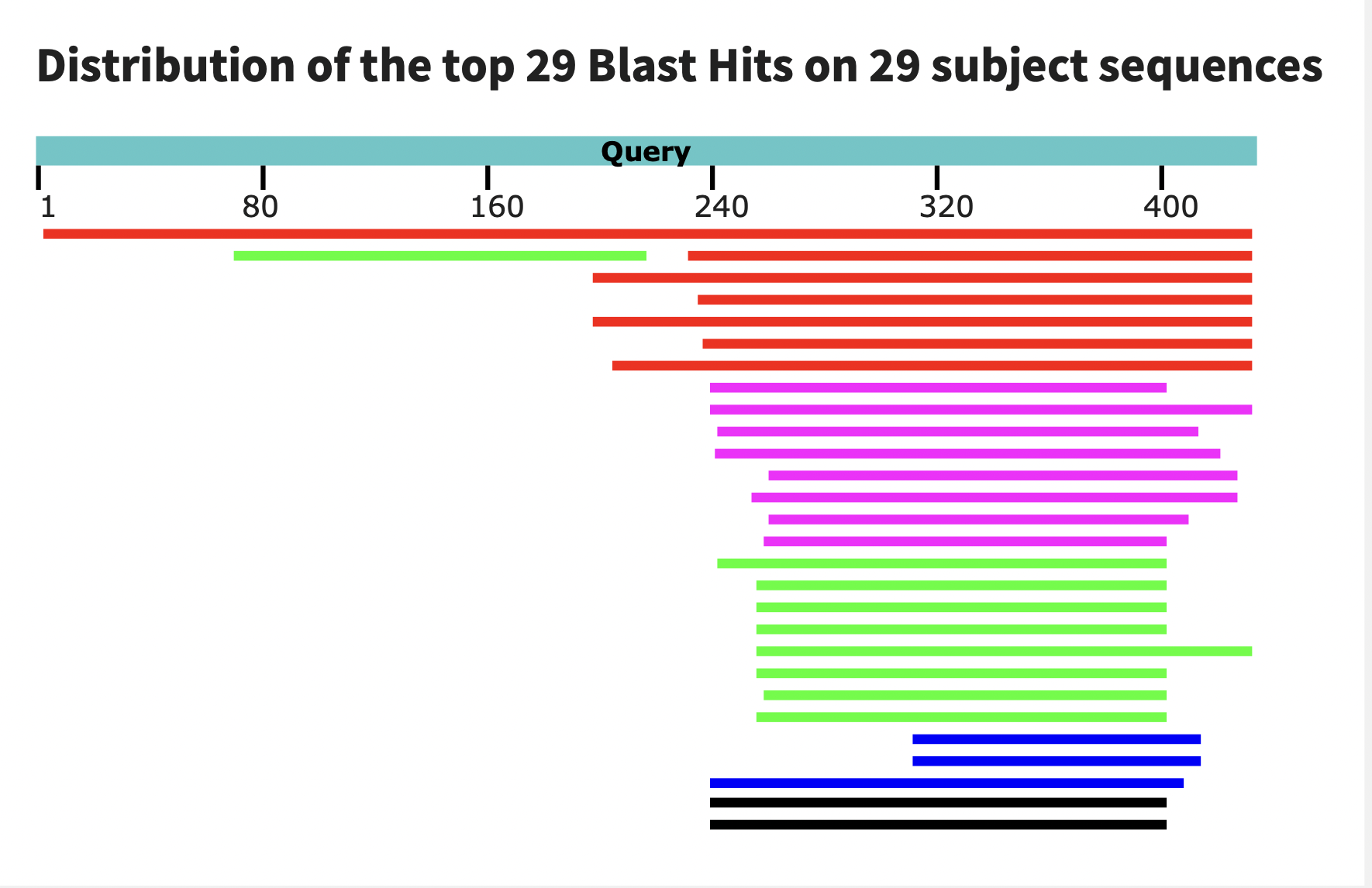
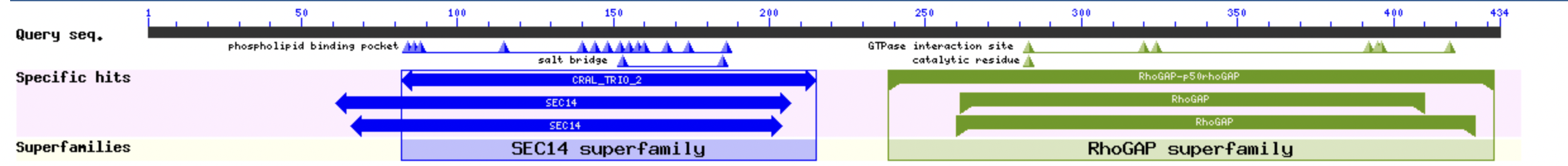
Всего Protein Blast предоставил 29 находок. Для каждой из них процент идентичности не был высок: 61,34% для лучшей находки (RhoA-GDP-Pi in Complex with RhoGAP человека), и примерно 52-56% идентичности для следующих 5 находок. У лучшей находки длина сопоставима с входной последовательностью (434 и 439 остатка), другие последовательности с идентичностью > 50% короче, порядка 200-240 а.о. Если посмотреть на выравнивание лучших находок на исходную последовательность (Рисунок 1), то можно заметить, что чаще выравнивается участок с 240 по 400 позиции относительно входной последовательности. Они соответствуют домену RhoGAP суперсемейства. (Рисунок 2) К этому домену принадлежит каталитический остаток и сайт взаимодействия с ГТФазой. На N-конце выделен домен, характерный для SEC14 суперсемейства. Для него показаны солевые мостики и карман, связывающий фосфолипиды.
Структура более консервативна, чем последовательность. Поэтому можно поработать с белком с идентичностью 61% под идентификатором PDB 6R3V.
Рисунок 1. Распределение выровненных участков из выдачи Protein Blast
Рисунок 2. Домены, показаны на основе лучших последовательностей по выравниванию в Protein Blast
Рассмотрение остатков
Для анализа были предложены следующие замены: N400E, R283K, K320E. Можно заметить, что эти замены ближе к C-концу белка, то есть должны попадать в домен RhoGAP.
На Рисунке 3 приведен фрагмент выравнивания, в который попадает домен RhoGAP. Это структура RhoA-GDP-Pi в комплексе с RhoGAP (белок человека) - нашим целевым белком. Эта структура разрешена методом рентгено-структурного анализа и имеет разрешение 1.75 Å, метрики качества довольно хорошие. В этой структуре нет модели для позиций 1-234 и структрура как раз начинается с домена RhoGAP, где и располагаются исследуемые остатки.
Рисунок 3. Фрагмент выравнивания целевого белка с лучшей находкой Blast. Соответствует RhoGAP домену. Красные прямоугольники - остатки, которые будут мутированы.
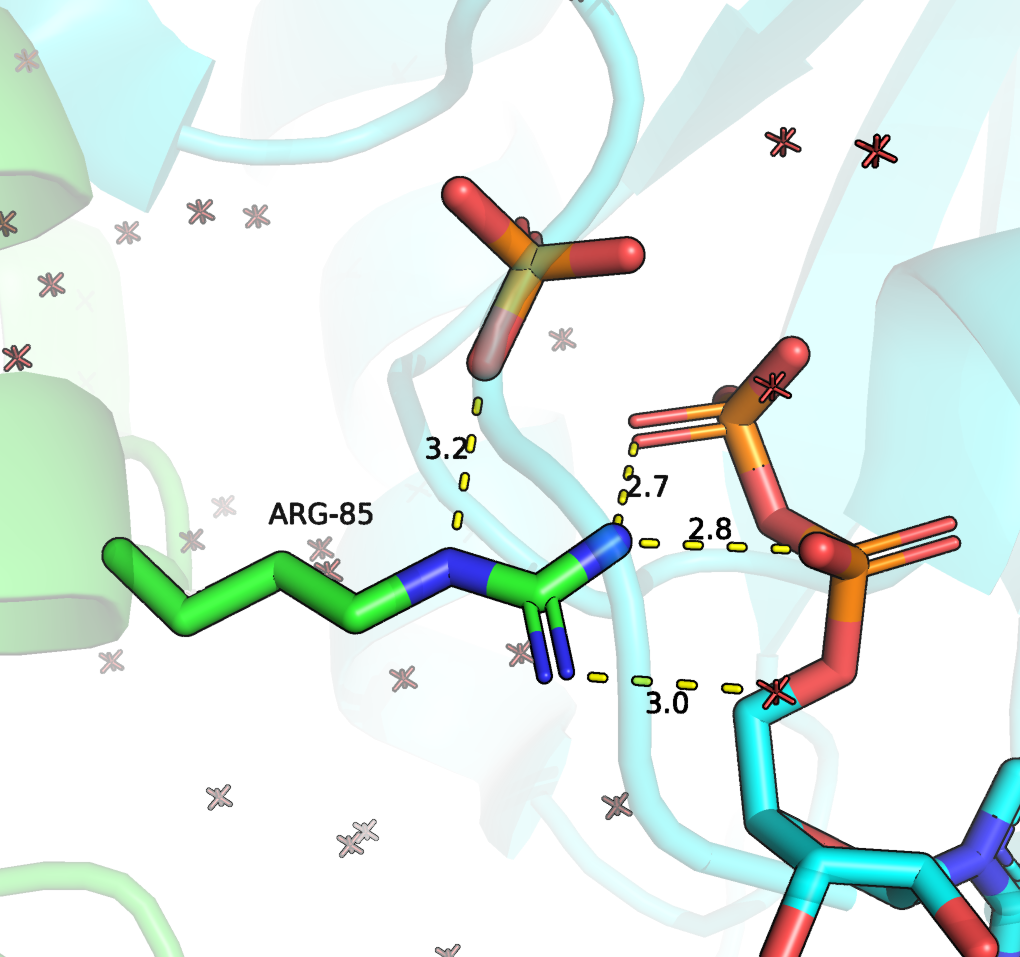
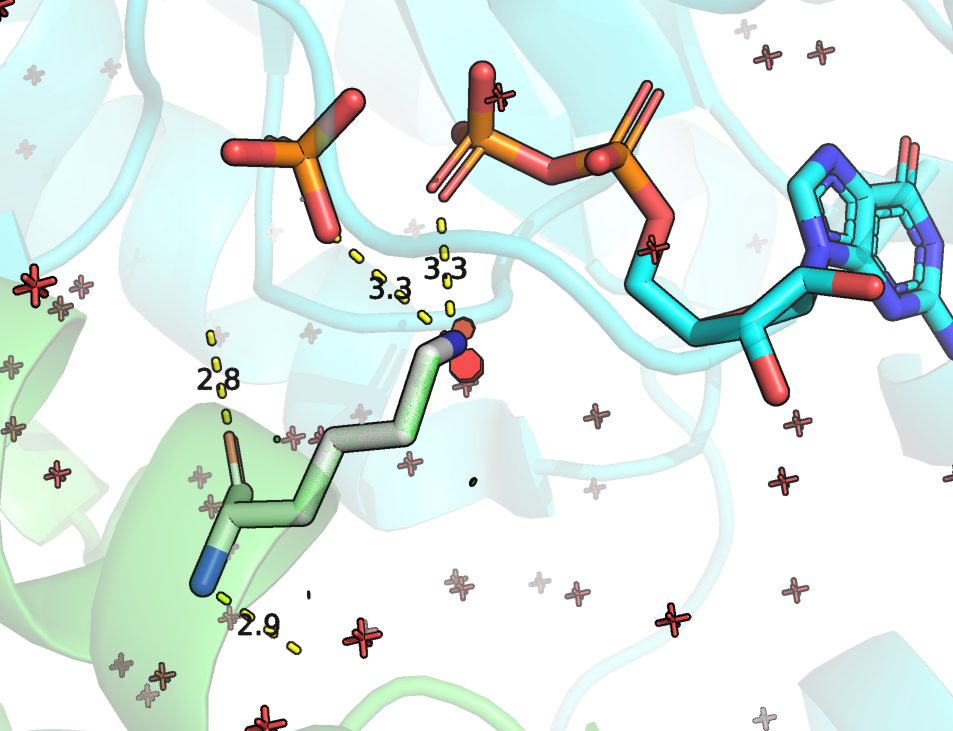
До моделирования замен посмотрим на положение остатков. Остаток ARG-283 без сомнения очень важен. Это и есть тот каталитический остаток, который приникает в активный сайт Rho ГТФ-азы и стабилизирует интермедиат. Он показан Рисунке 4, в данной структуре он имеет номер 85.
Рисунок 4. Положение ARG283 (в данной структуре имеет номер 85) и его взаимодействия с GDP. Зеленым цветом отмечена цепь RhoGAP домена, зеленым - ГТФазы.
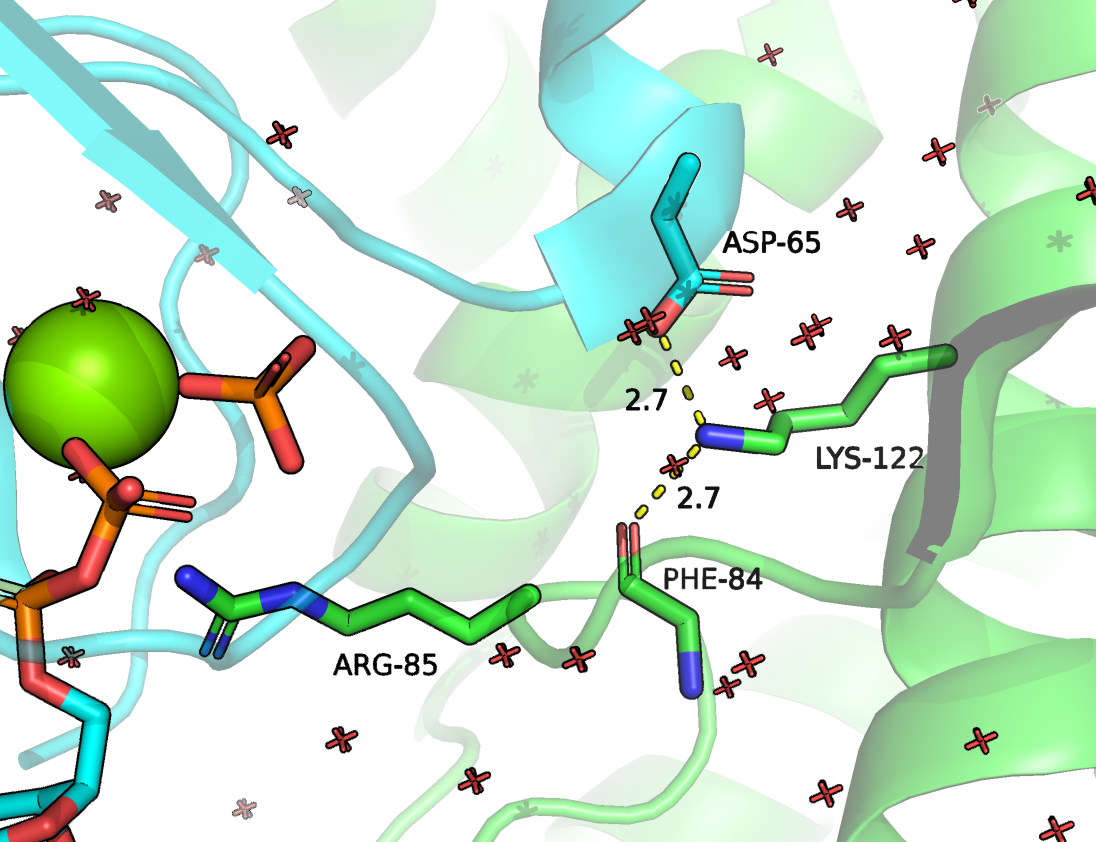
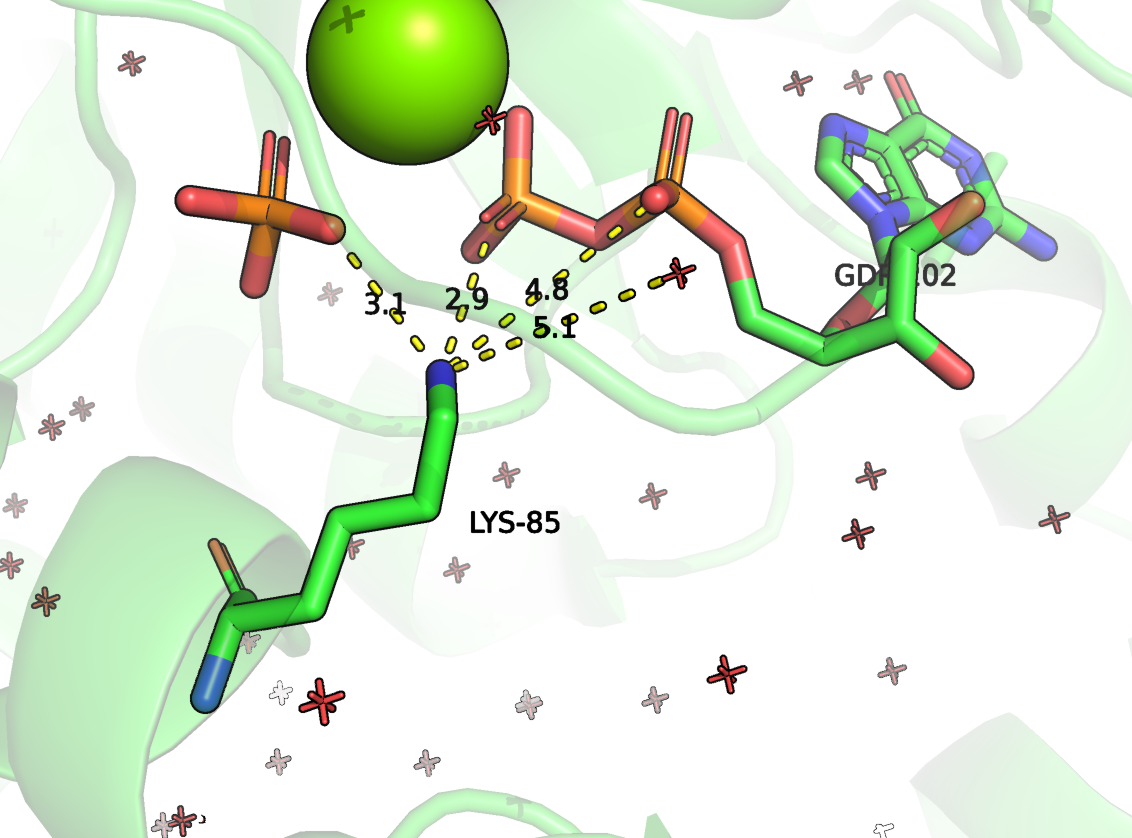
Рисунок 5. Положение ARG283 (в данной структуре имеет номер 85) и LYS320 (номер 122) и их контакты. Зеленым цветом отмечена цепь RhoGAP домена, зеленым - ГТФазы.
LYS-122 должен отвечать за взаимодействие между цепями, образуя солевой мостик с ASP-62 ГТФазы (Рисунок 5). Также должна быть важной водородная связь с кислородом остова PHE-84. Он находится в непосредственной близости к каталитическому остатку ARG-85. Наверняка LYS-122 вносит вклад в удерживании петли в правильном положении, чтобы ARG-85 мог проникнуть в активный сайт ГТФазы.
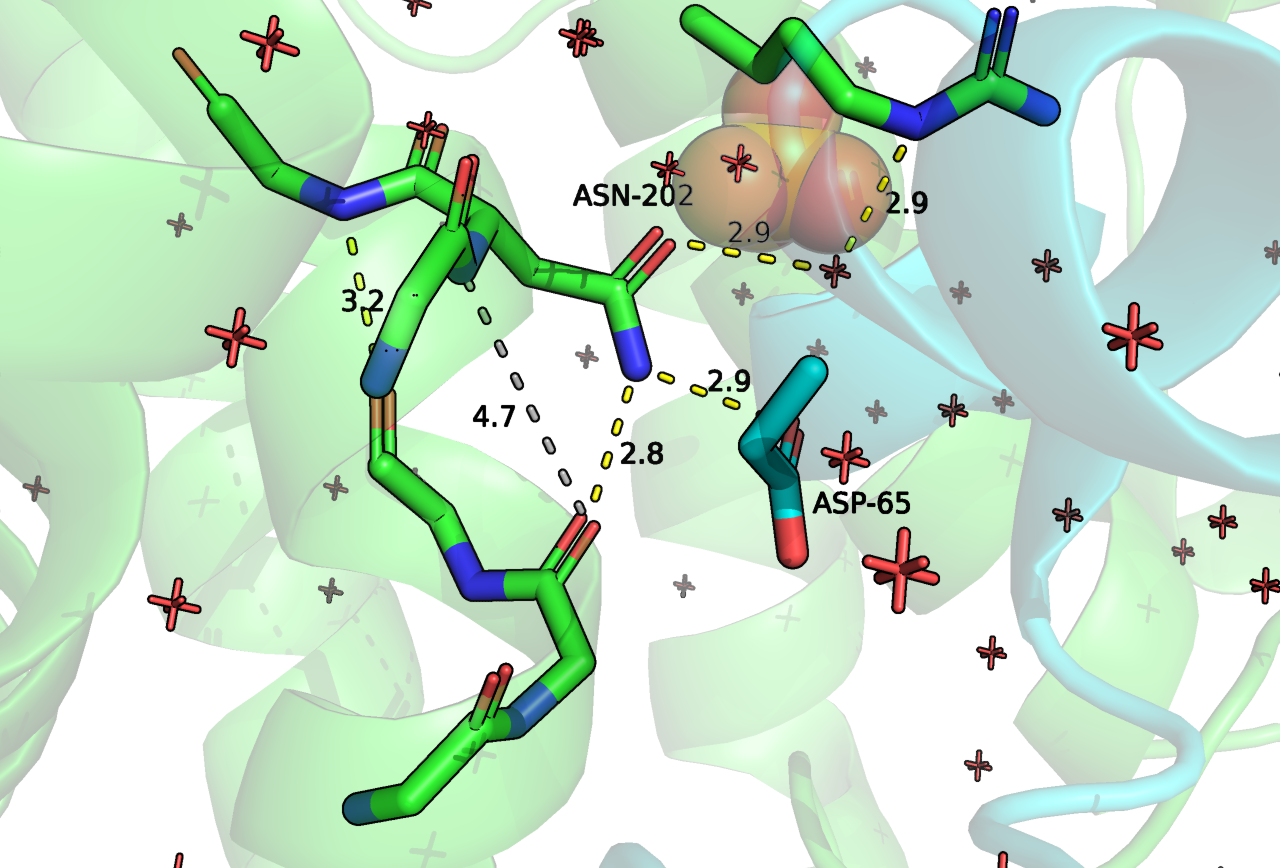
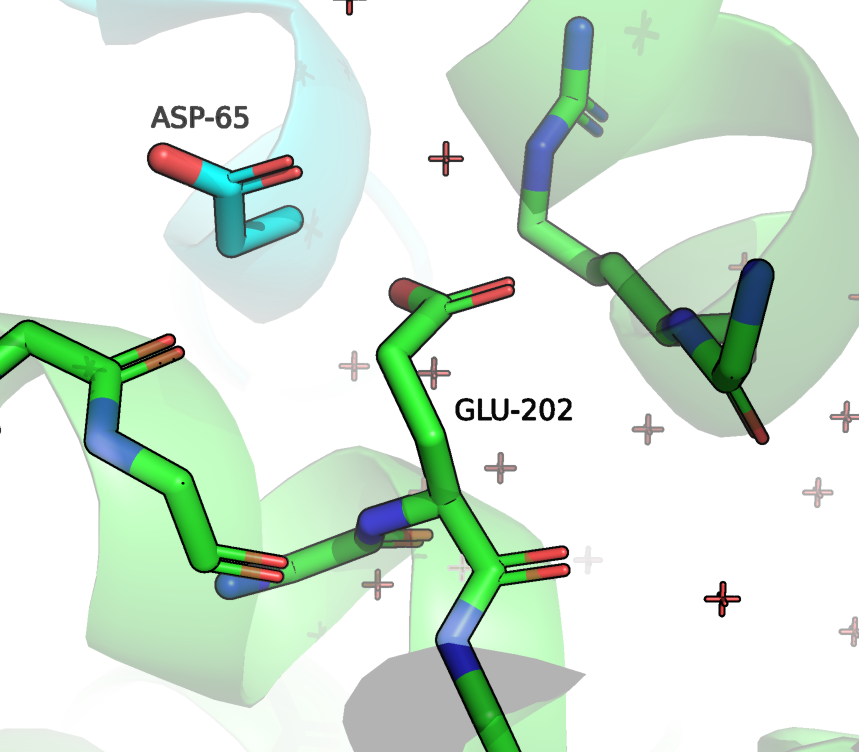
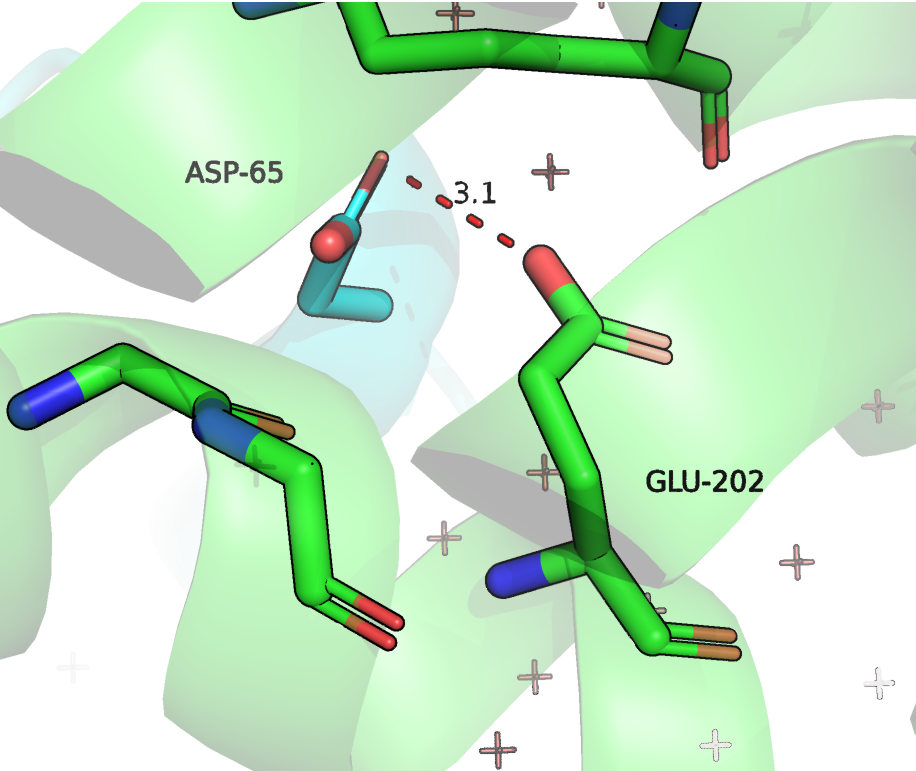
Что касается ASN-400 (202 на Рисунке 6), то он образует водородную связь с ASP-65. C ASP-65 мы уже встречались (Рисунок 4), это важный остаток для контакта двух белков. Кроме того, боковая группа ASN-202 образует водородную связь с остовом. В альфа спирали, которой принадлежит анализируемый остаток наблюдается нарушение хода (С-O [i] и N-H [i+4] слишком далеки), но спираль при этом сильно не теряет форму, полагаю, что как раз за счет взаимодействия с C-O [i] с ASN-202. Поэтому мне кажется, что этот остаток также важен для выполнения и структурной роли.
Рисунок 6. Положение и контакты ASN202 (в данной структуре имеет номер 85). Зеленым цветом отмечена цепь RhoGAP домена, зеленым - ГТФазы.
Мутации и их возможная функциональная роль
С помощью инструмента Pymol Mutagenesis я примерила новые остатки на данные позиции.
В первом случае каталитический ARG я пропробовала заменить на остаток LYS, тоже с заряженной боковой группой. Из возможных ротамеров был найден тот единственный, для которого сохранятся ионные связи с кислородами фосфатных групп. Strain для такого ротамера составляет 23.71, что является допустимым значением. При таком расположении все же образуется небольшой clash, а также расстояния в катионных мостиках значительно длиннее (3.3 против 2.7-2.8 ангстрем, которые наблюдались для ARG).
После применения инструмента sculpting удалось избежать перекрывания радиусов. Можно заметить, что даже при таком неплохом положении нельзя добиться такого же числа водородных связей, как в случае остатка ARG (Рисунок 4 и Рисунок 7). Не будет водородной связи с кислородом альфа-фосфата. Непонятно, что будет с молекулой воды в активном центре, и как повлияет смещение молекулы воды или отсутствие связи с ней. В данном случае я прогнозирую снижение активности фермента.
Рисунок 7. Замена ARG283 (85) на LYS320. Изображения до и после применения sculpting
Остаток GLU на месте ASN помещается уже хуже, значение strain пограничное - 25.98. Недалеко расположен ASP-65, и если расстояния между новым GLU и ASP-65 будут малы, то мы можем получить отталкивание. После скульптурирования действительно оказалось, что отрицательные заряды раположены близко в пространстве (Рисунок 8). К тому же остаток ASP-65 образует ионную связь с LYS-122 (Рисунок 5) и сильное смещение этого остатка может нарушить связь, что также может сказаться на взаимодействии RhoGAP и ГТФазы. Еще пропадет связь с кислородом остова, которая могла поддержтвать спираль (Рисунок 6).
У этих белков не очень большая площадь контакта. Я бы сказала, что такая замена могла бы повлиять на связывание и успешное попадание петли в активный центр ГТФазы.
Рисунок 8. Замена ASN400 (202) на GLU400. Изображения до и после применения sculpting
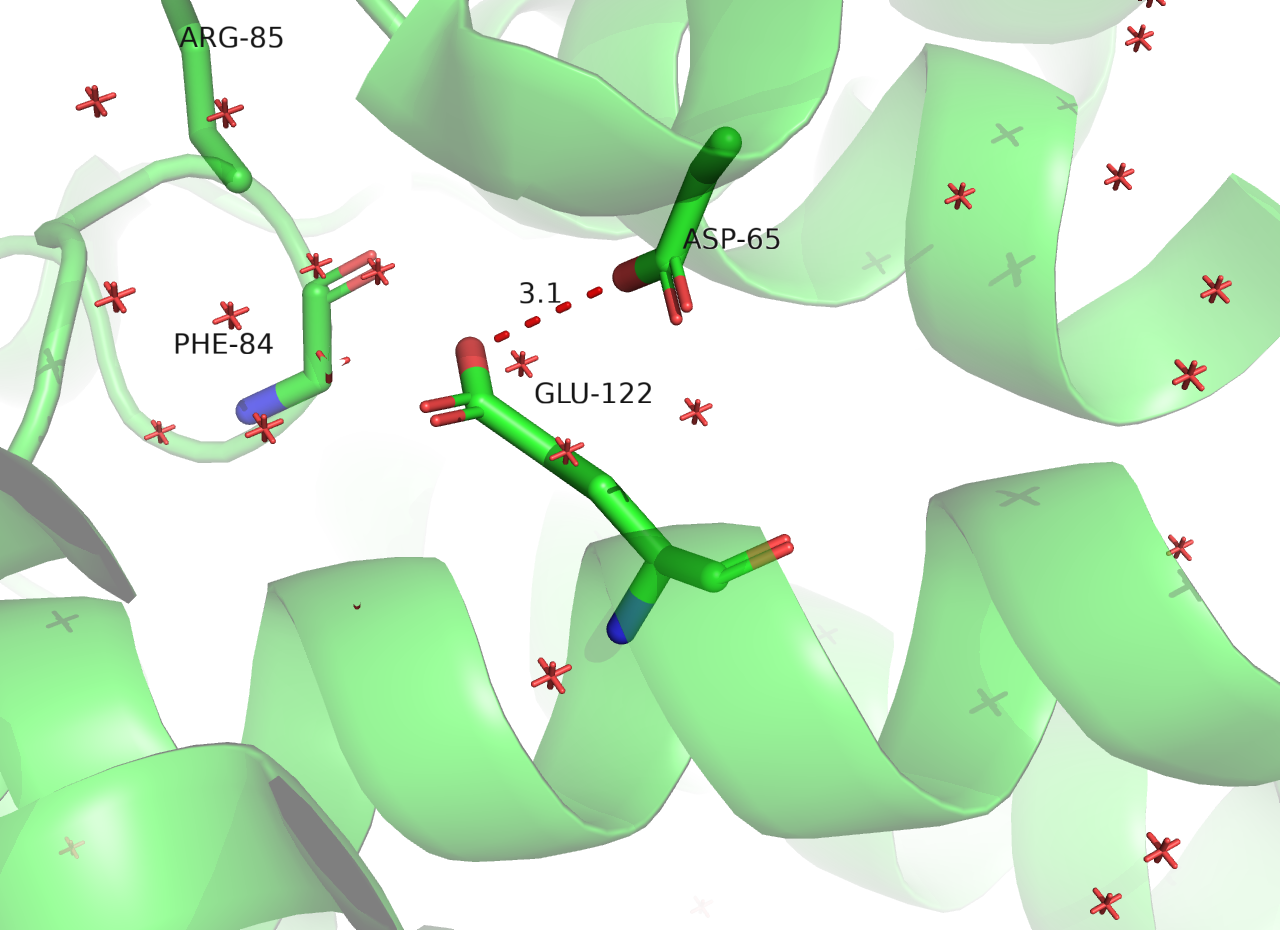
При замене на LYS-122 на GLU-122 не возникло перекрывания радиусов (strain = 16.53). После sculpting заряды GLU-122 и ASP-65 оказались на довольно близком расстоянии - 3.1 ангстрем. Кроме отталкивания, больше невозможна водородная связь с кислородом остова следующего за каталитическим остатка. Полагаю, что это положение петли станет менее жестким, молекула может стать менее функциональной.