A-, B- и Z-формы ДНК
Практикум №2
Получение моделей структур форм ДНК
С помощью программы fiber были получены структуры A-, B- и Z-форм ДНК. Структуры дуплексов вы можете найти в архиве.
Сравнение структур трёх форм ДНК с помощью средств JMol
С помощью программы JMol мною были визуально определены большая и малая бороздки, затем с помощью программы ACD/ChemSketch было получено изображение одного из азотистых оснований (тимина). Красным цветом на изображении выделены атомы, смотрящие в сторону большой бороздки, а синим – в сторону малой.
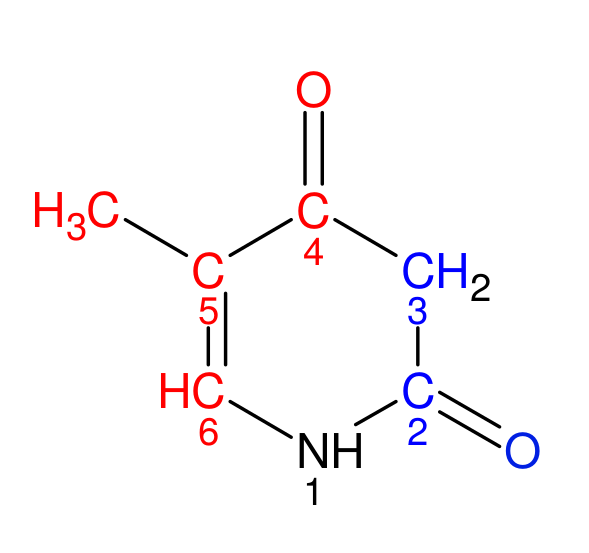
Рисунок 1. Структура тиминового азотистого основания
- В сторону большой бороздки обращены атомы: dt3.c4 or dt3.o4 or dt3.c5 or dt3.c6 or dt3.c7
- В сторону малой бороздки обращены атомы: dt3.c2 or dt3.o2 or dt3.n3 or dt3.n1
| A-форма | B-форма | Z-форма | |
| Тип спирали | Правая | Правая | Левая |
| Шаг спирали (Å) | 2,55 | 3,37 | 3,625 (7,25/димер) |
| Число оснований на виток | 11 | 10,5 (?) | 12 |
| Ширина большой бороздки (Å) | 16,6 (Adenosine) | 17,2 (Adenosine) | 7,2 (Guanosine) |
| Ширина малой бороздки (Å) | 8,0 (Adenosine) | 11,7 (Adenosine) | 15,2 (Guanosine) |
Таблица 1. Сравнение структур различных форм ДНК
Определение параметров структур нуклеиновых кислот
По средним значениям торсионных углов двухцепочечный участок структуры тРНК больше всего похож на B-форму.
| Обозначение | Значения (tRNA) | Значения (DNA) |
| α | -20.0 | -50.6 |
| β | 1.9 | 77.1 |
| γ | 69.3 | 38.7 |
| δ | 89.6 | 139.8 |
| ε* | -132.9 | |
| ζ | -72.8 | -80.3 |
| χ | -131.9 | -106.0 |
Таблица 2. Торсионные углы тРНК и ДНК
*Примечание: Рассмотрение среднего для углов ε в структуре ДНК (PDBID = 1LLR) в целом лишено смысла, так как большинство значений близки к 180°, отличие на +1° от, например, 179.2° влечет смену знака и результат -179.8°.
Наиболее деформированный нуклеотид в структуре с ДНК – Цепь II, цитозин №9, c тРНК – Цепь I, цитозин №18.
Следующие нуклеотиды образуют стебли во вторичной структуре тРНК:
| Акцепторный стебель | 5'-901-907-3' 5'-966-972-3' |
| D-стебель | 5'-910-912-3' 5'-923-925-3' |
| T-стебель | 5'-949-954-3' 5'-958-965-3' |
| Антикодоновый стебель | 5'-926-931-3' 5'-939-944-3' |
В структуре присутствует 9 неканонических пар. Несмотря на то, что некоторые из приведенных пар являются "каноническими", они отличаются от таковых пространственной структурой.
52 56 0 # 12 | ....>B:.954_:[..U]U-**--A[..A]:.958_:B<....
53 16 9 # 13 x ....>B:.955_:[..U]U-**+-G[..G]:.918_:B<....
35 31 0 # 14 | ....>B:.937_:[..A]A-**--U[..U]:.933_:B<....
36 30 0 # 15 | ....>B:.938_:[..U]U-**--U[..U]:.932_:B<....
38 28 0 # 17 | ....>B:.940_:[..C]C-*---G[..G]:.930_:B<....
42 24 0 # 21 | ....>B:.944_:[..C]C-*---A[..A]:.926_:B<....
12 43 0 # 25 | ....>B:.913_:[..A]A-**+-A[..A]:.945_:B<....
13 7 0 # 26 | ....>B:.914_:[..A]A-**--U[..U]:.908_:B<....
14 46 9 # 27 x ....>B:.915_:[..G]G-**+-C[..C]:.948_:B<....
Рисунок 2. Пример неканонической связи A—U
Также имеются три пары, не относящиеся к стеблям и стабилизирующие третичную структуру РНК.
52 56 0 # 12 | ....>B:.954_:[..U]U-**--A[..A]:.958_:B<....
53 16 9 # 13 x ....>B:.955_:[..U]U-**+-G[..G]:.918_:B<....
17 54 1 # 28 + ....>B:.919_:[..G]G-----C[..C]:.956_:B<....
© Arsenii Loginovskii, 2016-2018