1. Визуальный аналз моделирования самосборки липидного бислоя
trjconv -f b_md.xtc -s b_md.tpr -o b_pbc_1.pdb -skip 20
Сделаем так, чтобы при переходе с одной стенки ячейки на другую, молекулы переносились полностью
trjconv -f b_md.xtc -s b_md.tpr -o b_pbc_2.pdb -skip 20 -pbc mol
Если включить анимацию в файле b_pbc_2.pdb , видно как из хаотично разбросааных молекл липидов образуется мицелла. Образование бислоя примерно начинается с модели номер 24, которой соответствует время 11500 фемтосекунд после начала моделирования.
2. Анализ результатов
3. Определим площадь, занимаемую одним липидом. Для этого получим размер ячейки из траектории:
g_traj -f b_md.xtc -s b_md.tpr -ob box_1.xvg
В файле box_1.xvg содержатся размеры ячейки: в первой колонке - время, во второй - размер в нанометрах по оис X, в 3-ей - по Y, в 4-ой - по Z. Ось X - нормаль к поверхности бислоя.
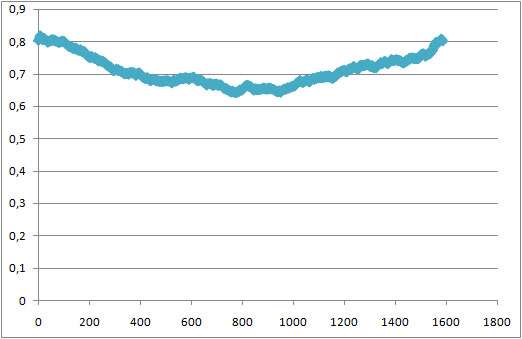
Всего 64 молекл липида, в одном слое - 32. График зависимости X*Y/32 от времени представлен ниже:
Во время начала образования мицеллы площадь одного липида минимальна, затем мицелла немного растянулась и площадь, занимаемая липидом возросла.
4.Определите изменение гидрофобной и гидрофильной поверхности в ходе самосборки:
g_sas -f b_md.xtc -s b_md.tpr -o sas_b.xvg
Файл sas_b.xvg содержит данные о гидрофильных и гидрофобных поверхностях, доступных растворителю. Изменение гидрофильной (красная) и гидрофобной (синяя) поверхностей во время моделирования:
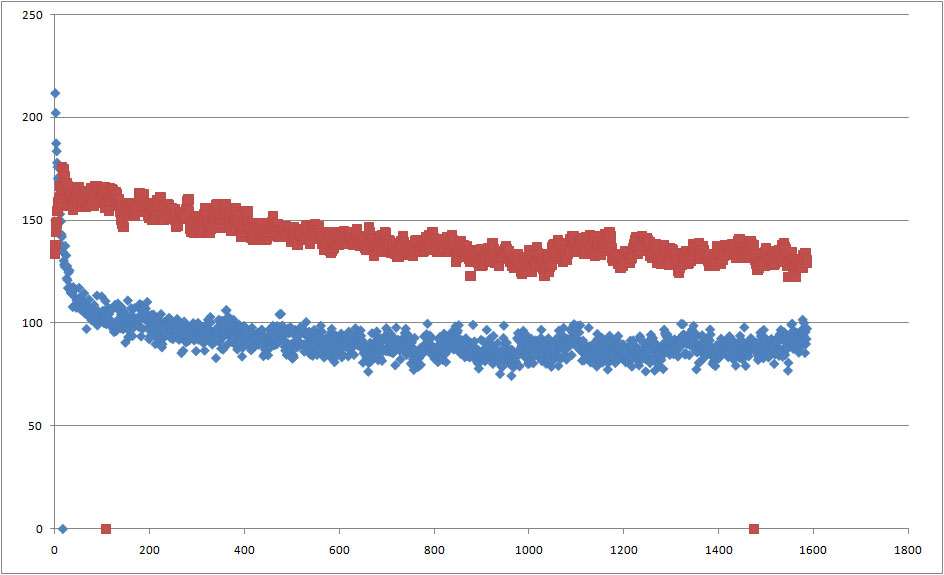
Для образования липидного бислоя главной движущей силой является уменьшение контакта гидрофобной части липида с растворителем. Как видно из рисунка уменьшается как гидрофильная так и гидрофобная поверхности, однако резкое уменьшение гидрофобной поверхности почему-то наблюдается почти сразу, еще до образования мицеллы.
5.Традиционной мерой оценки фазового состояния бифильных молекул является мера порядка. Для анализа нам понадобится индекс файл .
Для конца траектории:
g_order -s b_md -f b_md.xtc -o ord_end.xvg -n sn1.ndx -b 45000 -d X
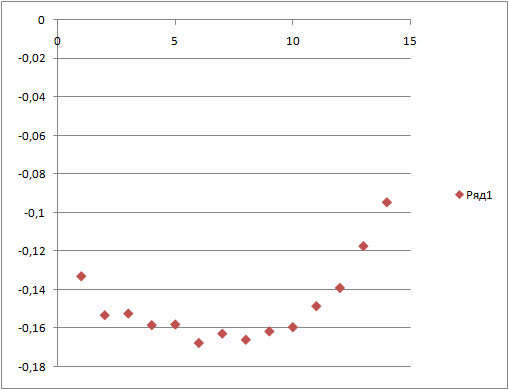
Для начала траектории:
g_order -s b_md -f b_md.xtc -o ord_start.xvg -n sn1.ndx -e 5000 -d X

Как видно из графиков, и в начале и в конце моделирования, атомы, расположенные ближе к середине липида, менее подвижны, чем головки и хвосты липида. В конце моделирования, когда сформировалась мицелла, подвижность всех атомов в целом ниже, чем в начале моделирования.