1.Основные возможности JalView
В данном задании необходимо было построить выравнивание 6 последовательностей гомологичных белков (по 2 из Архей, Эукариот и Бактерий) семейства HSP70. Я использовала белки с идентификаторами B2HPS1, A1UA31, Q6L0S7, Q97BG8, Q9LHA8, P22953. Для построения выравнивания использовалась программа JalView (а частности, Tcoffee). Раскраска производилась по схеме ClustalX с условием Identity Threshold = 100%. Выравнивание доступно по ссылке, рисунок 1 -полное выравнивание. Также на рисунке 2 отмечены примеры различных типов консервативаности: консервативных на 80% или более (C), абсолютно функционально консервативных (F), позиций с гэпами (G). Под функционально консервативными участками мы понимаем такие участки, где произошли замены аминоксилот, схожих по свойствам. Например, в позиции 169 у белка A1UA31 произошла замена аспарагиновой на глутаминовую кислоту. Эти аминокислоты относятся к отрицательно зараженным аминокислотам и похожи по своим свойствам. Другие примеры функционально консервативных замен: замена в 194 позиции у белка Q97BG8 лейцина на валин (гидрофобные аминокислоты) и замена в позиции 216 фенилаланина на лейцин у белков Q6L0S7 и Q97BG8.
 Рисунок 1. Выравнивание 6 гомологичных белков семейства HSP70
Рисунок 1. Выравнивание 6 гомологичных белков семейства HSP70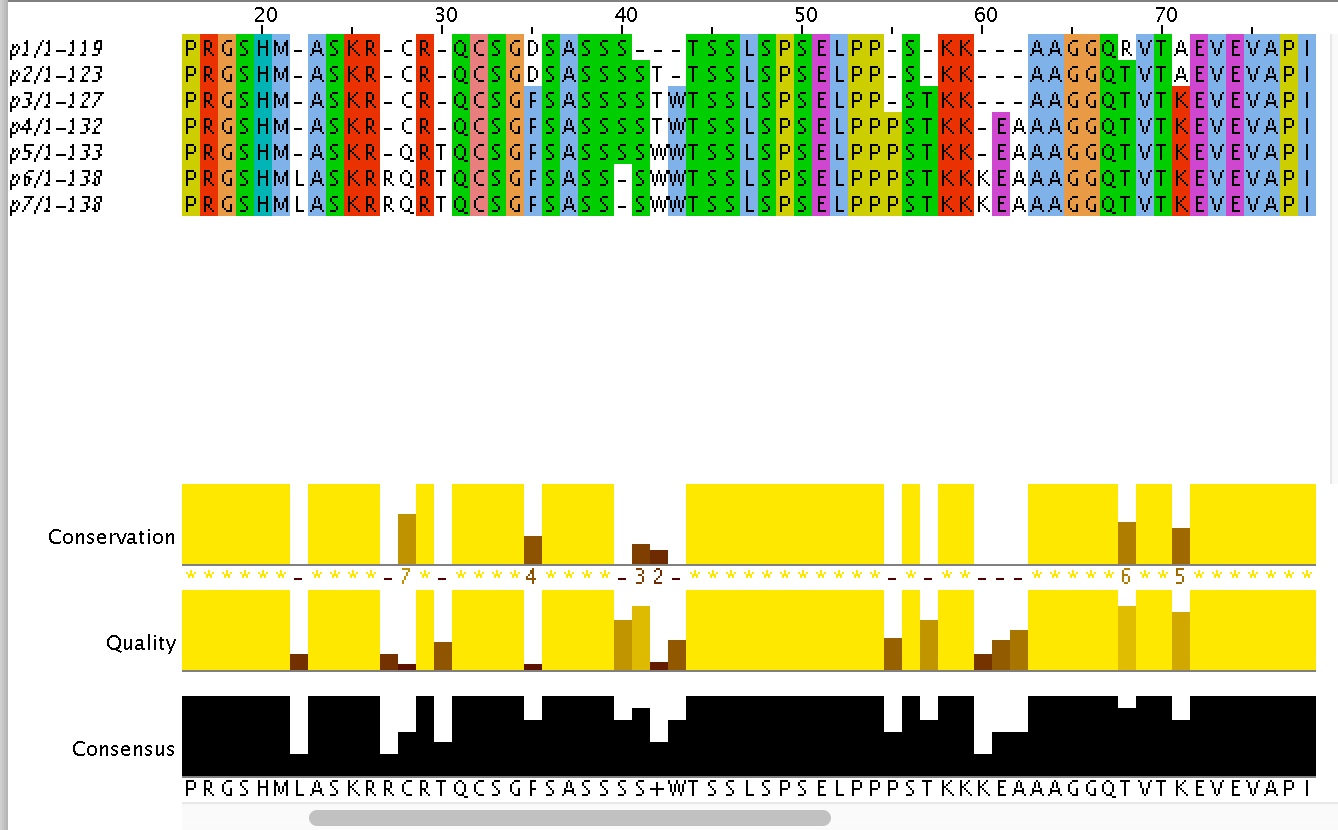 Рисунок 2. Участок выравнивания 6 гомологичных белков семейства HSP70
с разметкой
Рисунок 2. Участок выравнивания 6 гомологичных белков семейства HSP70
с разметкой | Параметр | Число | Процент |
| Абсолютно консервативные | 193 | 28,81% |
| Абсолютно функционально консервативные | 319 | 47,61% |
| Name | Sequence Lengtd | Aligned Length | Gaps | Gaps percent | Identity abs | Identity percent | Similarity abs | Similarity percent |
| A1UA31.1 | 621 | 670 | 49 | 7,31% | 345 | 51,49% | 23 | 3,43% |
| B2HPS1.1 | 621 | 670 | 49 | 7,31% | 347 | 51,79% | 20 | 2,99% |
| Q6L0S7.1 | 612 | 670 | 58 | 8,66% | 327 | 48,81% | 26 | 3,88% |
| Q97BG8.1 | 612 | 670 | 58 | 8,66% | 318 | 47,46% | 31 | 4,63% |
| Q9LHA8.1 | 649 | 670 | 21 | 3,13% | 313 | 46,72% | 32 | 4,78% |
| P22953.3 | 650 | 670 | 20 | 2,99% | 315 | 47,01% | 31 | 4,63% |
2.Небывалая эволюция последовательности белка
Для выполнения этого задания я взяла последовательность первых 119 аминоксилот белка 2J63 (цепь А). Затем с помощью команды msbar было создано 7 поколений белков с 7 сайтами мутаций на поколение. Ссылка на текста скрипта. Результат был записан в один файл, затем было построено выравнивание (ссылка на компьютерное выравнивание). В таблице 3 приведены первые 10 мутаций с указанием поколения, в котором они случались.| Номер позиции | Первоначальная аминокислота | Замененная аминокислота | На каком этапе эволюции |
| 1 | M | S | p3-p4 |
| 10 | H | N | p2-p3 |
| 14 | L | - | p5-p6 |
| 22 | - | L | p5-p6 |
| 27 | - | R | p5-p6 |
| 28 | C | Q | p4-p5 |
| 30 | - | T | p4-p5 |
| 35 | D | F | p2-p3 |
| 40 | - | S | p5-p6 |
| 43 | - | W | p2-p3 |
1)Гэп из позици 41 белка p2 необходимо поместить в позицию 43.
2) К в позиции 62 белков 1-3 нужно переместить в 59 позицию
3)В позициях 116 и 117 белков 6 и 7 необходимо поменять местами ГЭП и F, тогда между белками 4,5, 6,7 в позиции 117 будет идентичность.
4)В позициях 136 и 137 белков p4 и р5 необходимо поменять местами Y и гэп. Тогда в позиции 136 будет идентичность между белками 4,5,6,7.
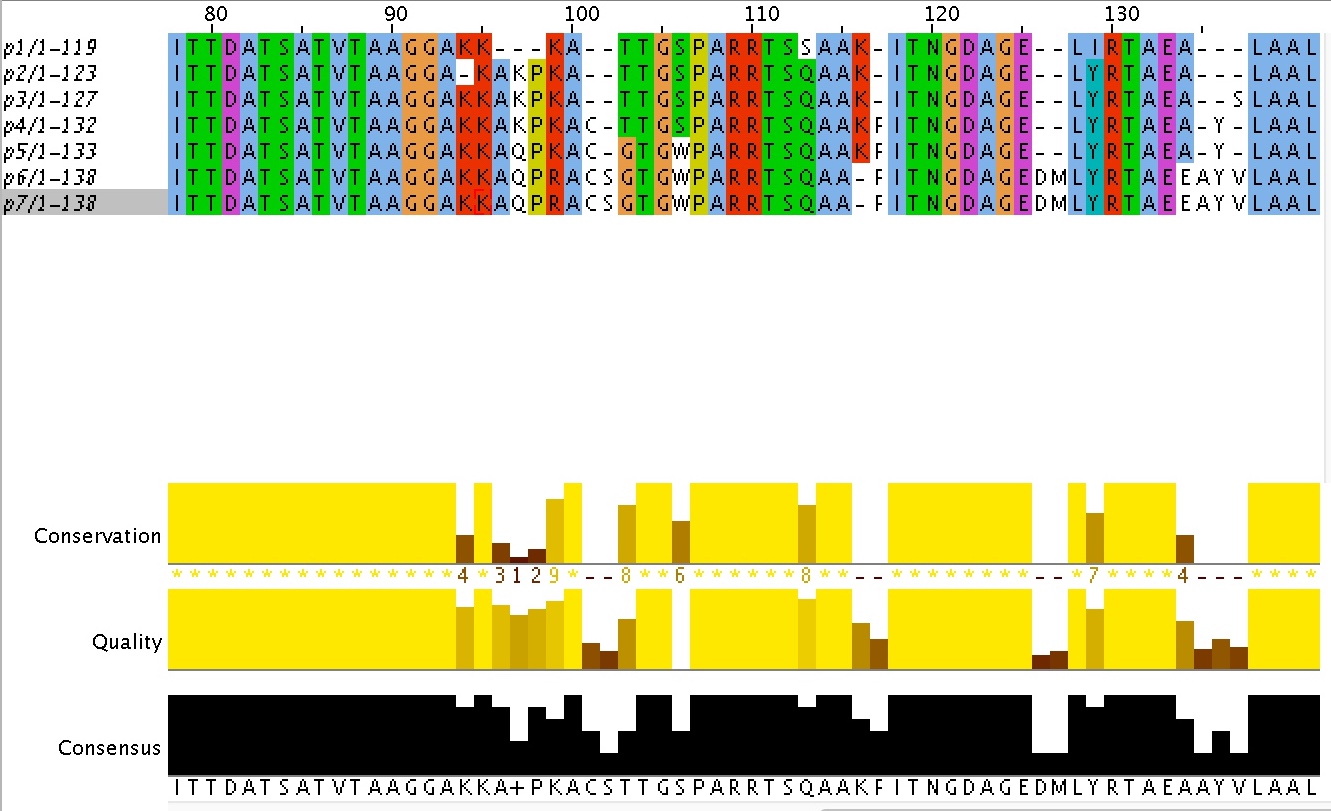 Рисунок 3. Участок исправленного вручную выравнивания между 7 поколениями белков, подверженных активному мутагенезу
Рисунок 3. Участок исправленного вручную выравнивания между 7 поколениями белков, подверженных активному мутагенезу
3.Мутирование на уровне нуклеотидной послеовательности
Для данного эксперимента я выбрала белок ANE84085.1 длиной в 76 аминоксилотных остатков из организма Bacillus cereus stdain A1, являющийся транскрипционным регулятором. Его кодирующую последовательность я взяла из полного генома данного штамма (идентификатор NZ_CP015727.1:17812-18039). С помощью той же комнады msbar был проведен аналогичный эксперимент с мутированием в 7 поколениях , перевод нуклеотидной последовательности в аминокислотную осуществлялся с помощью команды tdanseq. Ссылка на текста скрипта. Затем выравнивание из 2 и 3 задания я обработала с помощью команды infoalign, c параметром identity =70.0, то есть позиции, консервативные на более чем 70%. Результат можно увидеть в таблице 4. Белки 1-7 из второго задания, мутации на уровне а.к.о., 8-14 -из третьего задания, мутации на уровне н.п.| Name | Sequence Lengtd | Aligned Lengtd | Gaps | Identity abs | Identity percent | Similarity abs | Similarity percent |
| p1_1-119 | 119 | 141 | 11 | 109 | 77,30% | 1 | 0,71% |
| p2_1-123 | 123 | 141 | 11 | 114 | 80,85% | 1 | 0,71% |
| p3_1-127 | 127 | 141 | 9 | 120 | 85,11% | 0 | 0% |
| p4_1-132 | 132 | 141 | 8 | 129 | 91,49% | 0 | 0% |
| p5_1-133 | 133 | 141 | 7 | 127 | 90,07% | 0 | 0% |
| p6_1-138 | 138 | 141 | 3 | 122 | 86,52% | 1 | 0,71% |
| p7_1-138 | 138 | 141 | 3 | 122 | 86,52% | 1 | 0,71% |
| p8_1-76 | 76 | 108 | 18 | 23 | 21,30% | 3 | 2,78% |
| p9_1-83 | 79 | 108 | 17 | 38 | 35,19% | 0 | 0% |
| p10_1-83 | 79 | 108 | 17 | 36 | 33,33% | 2 | 1,85% |
| p11_1-89 | 82 | 108 | 16 | 37 | 34,26% | 0 | 0% |
| p12_1-95 | 91 | 108 | 13 | 25 | 23,15% | 4 | 3,70% |
| p13_1-99 | 93 | 108 | 9 | 31 | 28,70% | 2 | 1,85% |
| p14_1-101 | 95 | 108 | 9 | 24 | 22,22% | 3 | 2,78% |
4.Take home messages
1) "Эволюционирует нуклеотидная последовательность генома". Как правило, эволюционные процессы работают на уровне организма (если появившийся в результате мутации нуклеотидной последовательности признак окажется полезным в той среде обитания, где проживает организм, то организм сумеет оставить больше потомков и передаст им этот признак, который таким образом закрепится в популяции). То есть действию отбора подвергаются белки и их функционирование в организме или в популяции (популяционный отбор)2)"Только мутации в половых клетках наследуются". В современной биологии развиволось направление так называемой эпигенетики [1], науки, изучающей изменение фенотипа, вызванные механизмами, отличными от изменений последовательности ДНК, таких как метилирование ДНК, деацитилирование гистонов и т.д. Ярким примером влияния эпигенетического наследования является наследование закручиваемости спирали раковины у прудовиков [2].
3)"В гомологичных последовательностях живущих сегодня организмов мы видим почти исключительно мутации, прошедшие отбор". У эукариот геном состоит из кодирующих участков (экзонов) и некодирующих (интронов), затем в процессе сплайсинга [3] из преМРНК вырезаются интроны и трансляция происходит уже только с экзонов. Поэтому интроны, как правило (за исключением сайтов сплайсинга, крайних участков), не подвергаются действию отбора и там накапливаются мутации. Поэтому филогенетику близкородственных видов строят на основании нуклеотидных последовательностей интронов.
4) "Последовательность белка обычно под стабилизирующем отбором, т.е. отбор действует против мутаций а.к.о. " Контрпример: серповидноклеточная анемия [4]. В гемоглобине происходит замена 1 глутаминовой кислоты на валин, в результате этого гемоглобин не может принять правильную конформацию и нормально выполнять свои функции. В гомозиготном состоянии данная мутация летальна, но в гетерозиготном она закрепилась в популяции, тк люди, имеющие такую мутацию устойчивы к малярийному плазмодию. Также отбор в легких цепях антител тоже не стабилизирующий, а похож на движущий, специальными ферментами вносят мутации в последовательность ДНК, что позволяет увеличить их вариабельность в различных организмах (этот пример может считаться контрпримером на утверждение, что мутации происходят случайно. Как видно из представленного примера, так происхоит не всегда.).
5)"Гомологичность последовательностей белков и их а.к.о. можно предсказать по высокому сходству фрагментов в блоках выравнивания. Для белков есть проверка: сходство структур" Это утверждение не всегда верно, так как в эволюции вторичных структур белков наблюдается консервативность каких-то элементов и блоков. Например, белки, выполняющие одинаковые функции, но не произошедшии от одного предка, имеют похожую вторичную структуру. Это явление в биологии называется аналогия [5]. А похожесть во вторичной структуре определяется наличием функционально консервативных позиций, обеспечивающих сходство фрагментов блоков выравнивания.
6) "Универсальных границ сходства, свидетельствующего о гомологии, не существует". Возможно, универсальных границ и не существует, но существуют критерии гомологии (Первоначальные, по Ремане, положения, спец качества, переходных форм и дополнительные, такие как критерий состава, развития и гентический критерий [5]). Если объекты удовлетворяют большинству из этих критериев, то мы вправе считать их гомологичными.
7) Утверждение Вентера “So it's tde first living self-replicating cell tdat we have on tde planet whose DNA was made chemically and designed in tde computer. So it has no genetic ancestors. Its parent is a computer.” Конечно, их заслуга в синтезе ддлинной цепочки ДНК в несколько миллионов пар нуклеотиов неумалима, но все-таки они не сами сконструировали последовательность букв ДНК, а взяли готовый геном, возможно как-то подредактировав. Так что я не согласна с утверждением, что родителем этого генома является компьютер, ведь он использует готовые модели нуклеотидных послеователньостей, кодирующих достоверно работающие белки, прошедшие отбор и закрепившиеся в популяции.
8)"Локальные мутации накапливаются со временем: больше времени – больше мутаций – больше различий между потомками" Не всегда большая разница между какими-то признаками означает малое сродство. Например, у двугорбых верблюдов эритроциты имеют овальную форму, что позволяет сохранять крови нормальную тягучесть, но разница по этим генам между ними и другими копытными примерна равна разнице их с другими млекопитающими, хотя верблюды достоверно ближе к копытным. Кроме того в эволюции есть тенденция активных периодов нового видообразования (например, при освоении новых мест обитания или после возникновения важной апоморфии), таких как Кембрийский взрыв, и не всегда "молеклярные часы" "отсчитывают" время линейно. Например, Сарич установил, что различия между альбуминами мыши и крысы на порядок выше, чем между таковыми у человека и шимпанзе, а значит, молекулярные часы у грызунов идут быстрее [7].
5. Список литературы
[1] https://ru.wikipedia.org/wiki/Эпигенетика
[2] http://elementy.ru/problems/529
[3] https://ru.wikipedia.org/wiki/Сплайсинг
[4] https://ru.wikipedia.org/wiki/Серповидноклеточная_анемия
[5] https://ru.wikipedia.org/wiki/Аналогичные_органы
[6] https://ru.wikipedia.org/wiki/Гомология
[7] http://www.medicalbrain.ru/genetika/geny-belki-i-molekulyarnye-chasy.html