Структура тРНК и ДНК-белковые контакты
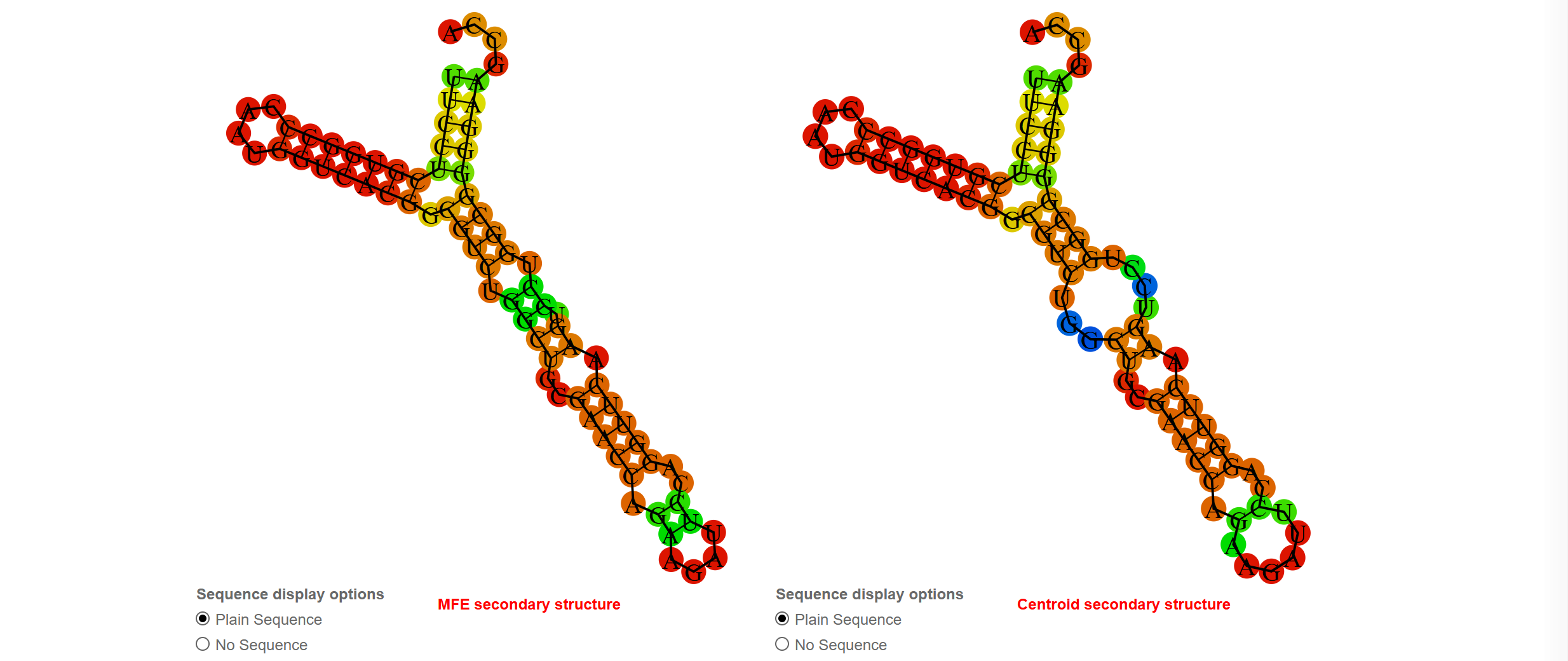
Предсказание вторичной структуры тРНК 1F7U
Как понятно из названия, TRNA(ARG) - это транспортная РНК для аргинина.
Из таблицы 1 видно, что einverted угадал только акцепторный стебель
| Участок структуры | Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера, Vienna |
| Акцепторный стебель | 5’- 1-7 -3’ 5’- 66-72 -3’ |
5’- 2-7 -3’ 5’- 66-72 -3’ |
5’- 1-5 -3’ 5’- 68-72 -3’ |
| D-стебель | 5’- 10-15 -3’ 5’- 22-25 -3’ |
- | 5’- 6-11 -3’ 5’- 17-23 -3’ |
| Антикодоновый стебель | 5’- 39-44 -3’ 5’- 26-31 -3’ |
- | 5’- 36-43 -3’ 5’- 25-28 -3’ |
| T-стебель | 5’- 48-56 -3’ 5’- 58-65 -3’ |
- | 5’- 49-55 -3’ 5’- 64-67 -3’ |
| Общее число канонических пар нуклеотидов | 27 | 7 | 27 |
.png)

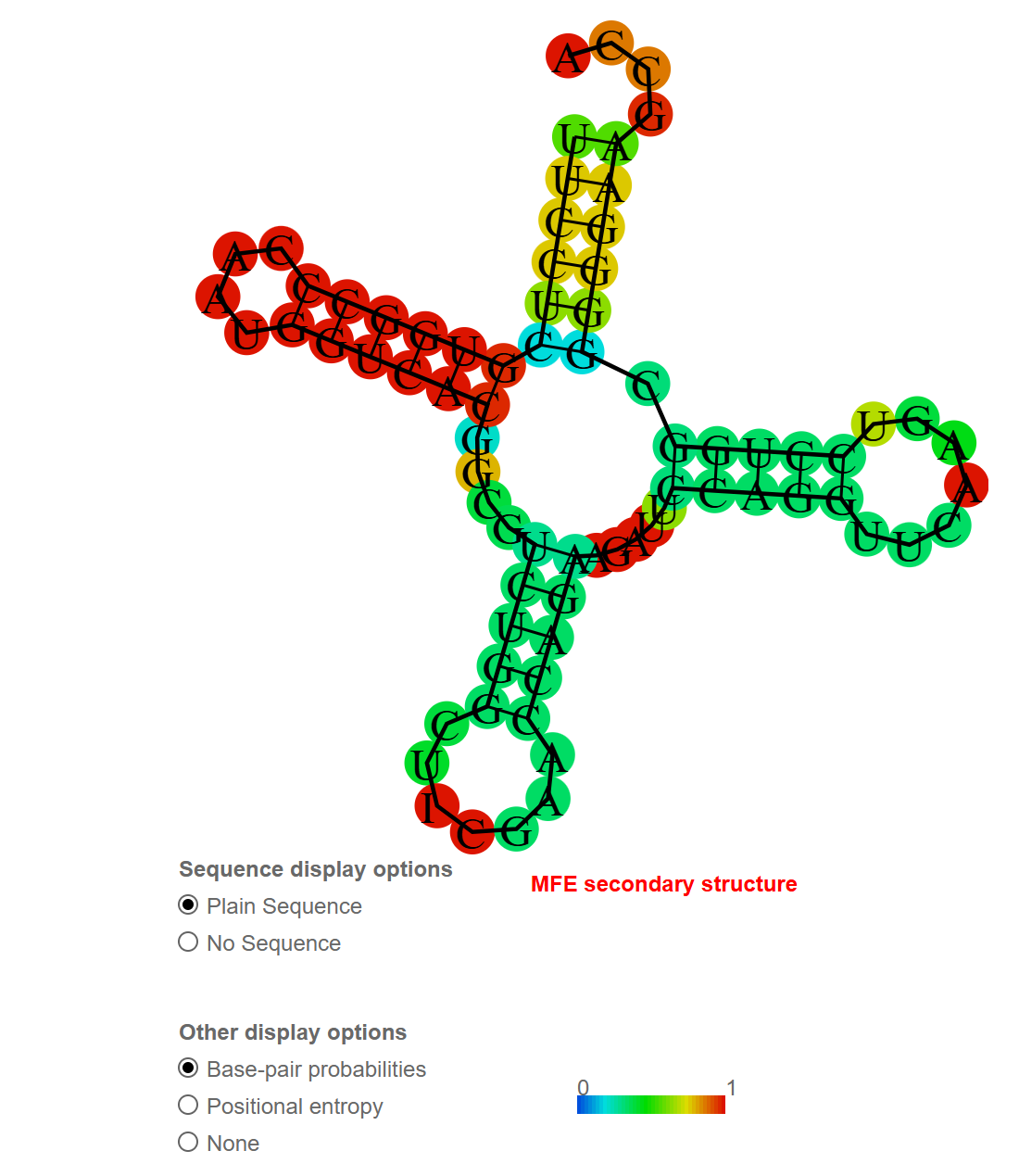
Выдача RNAfold:
1F7U_1|Chain A[auth B]|TRNA(ARG)|null UUCCUCGUGGCCCAAUGGUCACGGCGUCUGGCUGCGAACCAGAAGAUUCCAGGUUCAAGUCCUGGCGGGGAAGCCA ((((((((((((....))))))).((((.((((..(((((.((....))..))))).)).)).))))))))).... (-27.60)ДНК-белковые контакты
Скрипт для выделения групп в Jmol
Скрипт для отображения контактов для PyMol
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 126 | 69 | 195 |
| остатками фосфорной кислоты | 170 | 48 | 218 |
| остатками азотистых оснований со стороны большой бороздки | 105 | 11 | 116 |
| остатками азотистых оснований со стороны малой бороздки | 12 | 0 | 12 |
| остатками азотистых оснований (неуточнённые по бороздке) | 1 | 24 | 25 |
По данным таблицы наглядно видно преобладание контактов белка с азотистыми основаниями со стороны большой бороздки по сравнению с малой бороздкой. Это объясняется тем, что в большой бороздке расстояние между сахарофосфатными остовами ДНК больше, поэтому азотистые основания становятся более доступными для взаимодействия с аминокислотными остатками белка. В малой бороздке основания больше экранированы остовом ДНК, что затрудняет возможность прямых контактов. Также видно, что белок образует большее число контактов с фосфатными группами, чем с полярными атомами нуклеозида. Это связано с тем, что белок взаимодействует с сахарофосфатным остовом, удерживая комплекс белок–ДНК. Фосфатные группы несут отрицательный заряд, поэтому они эффективно взаимодействуют с положительно заряженными аминокислотами (аргинин и лизин например).
Для нашего комплекса nuclot выдаёт Segmentation fault, поэтому мы взяли струкуру 1I3J.

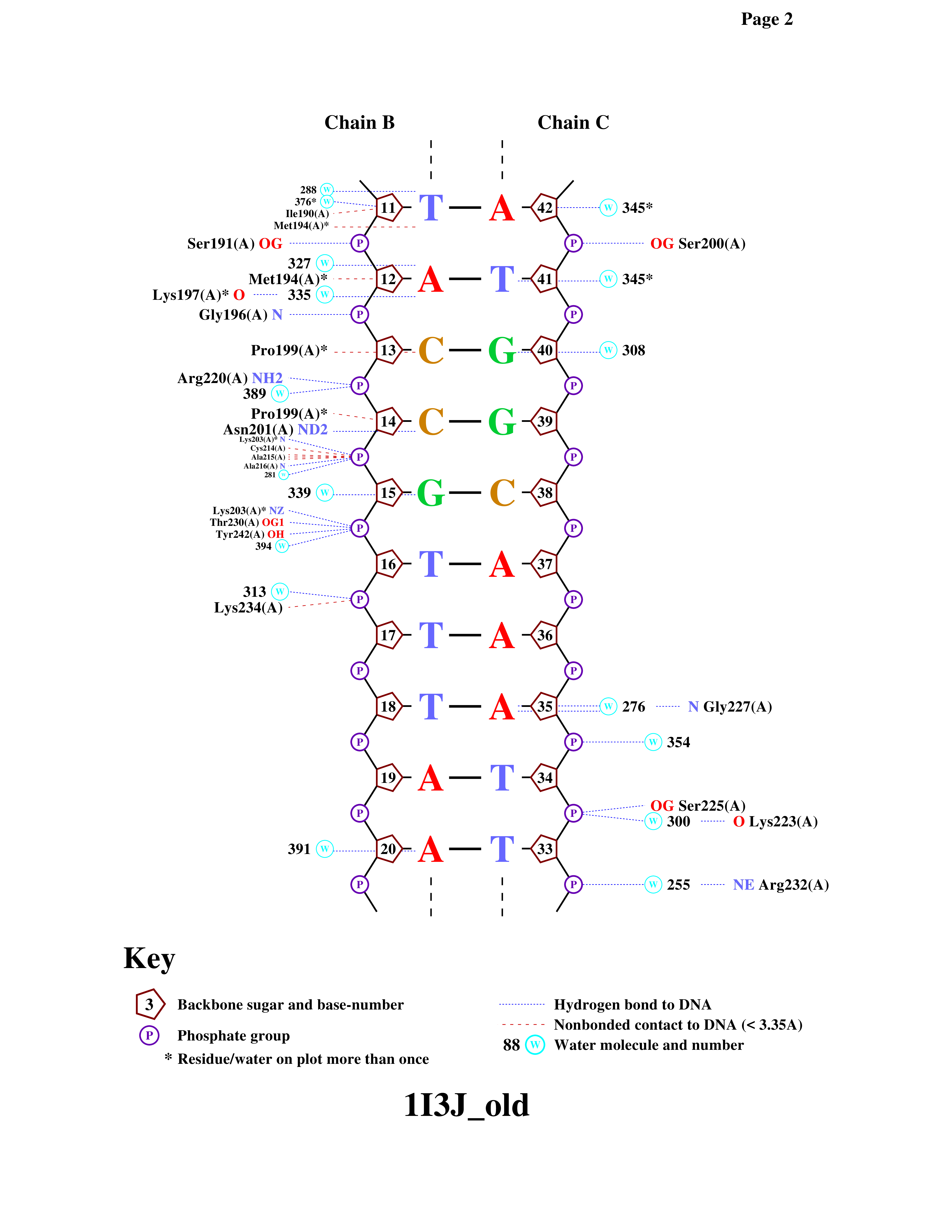
.png)
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 45 | 148 | 193 |
| остатками фосфорной кислоты | 59 | 27 | 86 |
| остатками азотистых оснований со стороны большой бороздки | 41 | 4 | 45 |
| остатками азотистых оснований со стороны малой бороздки | 9 | 0 | 9 |
| остатками азотистых оснований (неуточнённые по бороздке) | 0 | 18 | 18 |
Больше всего контактов образует Arg168, вот например с кислородом в сахаре аденина 49:
.png)
В целом, вероятно, это самый важный остаток в структуре потому что он образует контакты с двумя соседними аденинами, с DA-48 непосредственно с азотистым основанием:
