| Главная страница | Обучение | Обо мне | Ссылки |
Ферменты и метаболические пути. База данных KEGG. | |||
|
Задание 1. KEGG ORTHOLOGY Для работы был выбран метаболический путь биосинтеза лизина, а в нем была взята реакция, катализируемая двумя рядами ортологичных рядов: K05824 и K10978.
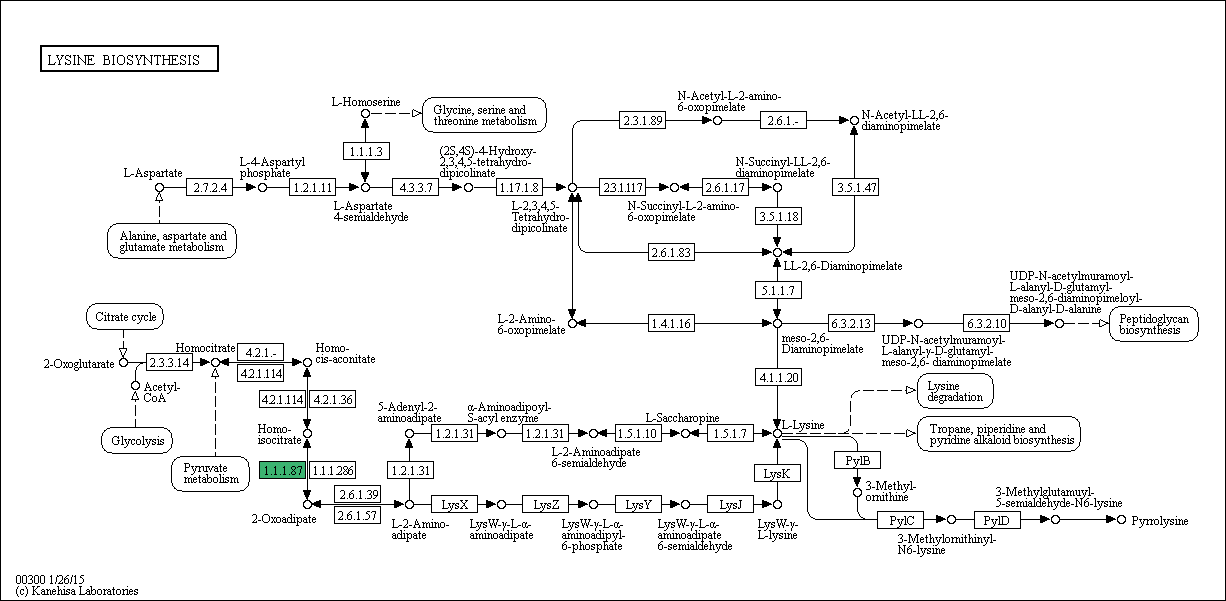
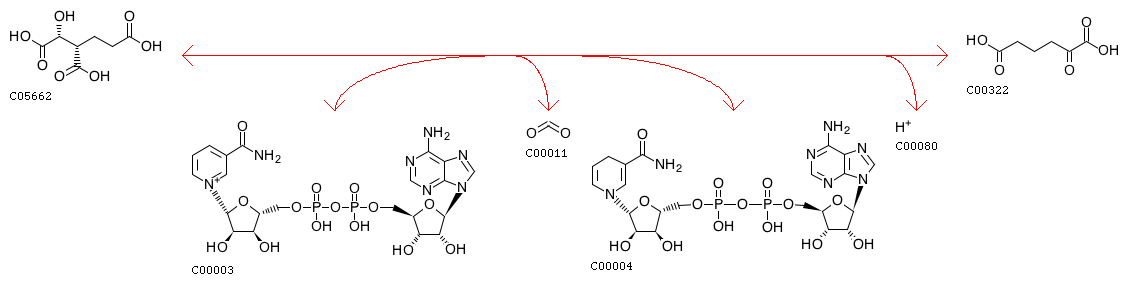
Это реакция образования 2-оксоадипата из гомоизоцитрата, имеющая номер 1.1.1.87 (отмечена зеленым на рис. 1). Ее схема приведена на рис. 2 (идентификатор реакции - R01934). Соответственно, ферменты, катализирующие ее, называются гомоизоцитратдегидрогеназами.
Рис. 1. Метаболический путь биосинтеза лизина (карта 00300). Зеленым выделена реакция, ферменты которой рассматриваются в данном практикуме.
Ряд К05824 содержит 123 белка (fasta-файл с этими белками), а К10978 - 69 белков (fasta-файл с этими белками). Затем в Jalview-проекте было построено множественное выравнивание этих двух ортологических рядов с помощью сервиса Muscle: ссылка на выравнивание в Jalview, исходные последовательности представлены в окне "input", выравнивание - в окне "alignment", проверенное и исправленное выравнивание, по которому затем строилось дерево - в окне "checked_alignment". Гомологичность белков в выравнивании В целом можно сказать, что множественное выравнивание получилось хорошее, так как белки явно гомологичны внутри ряда, и между рядами гомологичность тоже видна, так как есть достаточно большие (самый большой ~ чуть меньше 100 колонок) блоки консервативных позиций для всех (почти всех) белков выравнивания. При этом консервативность наблюдается не только по часто встречающимся аминокислотным остаткам вроде лейцина или валина, но и по более редким и, вероятно, функциональным остаткам (заряженным или образующим водородные связи, например). Проверка выравнивания В выравнивании присутствуют и такие белки, из-за которых возникают большие гэповые участки, однако в консервативных позициях они выровнены хорошо. Заметно, что у белков ортологического ряда K05824 в участки между блоками попадает большее количество аминокислот, чем у ряда К10978 (то есть белки К10978 в целом короче). Эти участки различий как раз должны использоваться для построения дерева, так как они индивидуальны (и могут объединяться в разные группы по степени схожести между собой). Также есть пара очень коротких белков, у которых возникает больше количество гэпов внутри блоков. Такие белки были удалены из выравнивания до построения дерева (последовательности A5DKS4_PICGU|K05824 и A5DKS5_PICGU|K05824). Построение филогенетического дерева Файл, на основе которого было построено дерево: checked.fasta. Использовался метод Neighbor-Joining с бутстреп-анализом (100 реплик). Дерево приведено на рис. 3. Рис. 3. Дерево гомоизоцитратдегидрогеназ из ортологических рядов К05824 и К10978. К сожалению, в данном дереве деление на клады не соответствует разделению белков на ортологические ряды, вернее, отделение части ортологического ряда К05824 от полного ряда К10978 происходит не в начале дерева. К тому же, оно поддержано низким количеством бутстреп-реплик (19). Однако первое разделение ветвей (которое могло бы быть разделением на два ряда) также поддержано низким процентом реплик (31). Тривиальных ветвей с резко отличающимися длинами нет. В целом, можно сказать, что эти два ряда очень хорошо выровнены между собой, поэтому Neighbor-Joining не смог разделить их на две клады. На страницу четвертого семестра | |||
| © Alexandra Boyko, 2014. Faculty of Bioengineering and Bioinformatics, MSU. | |||