Поиск в базах данных
Тема 4. ДНК метилтрансферазы млекопитающих.
ДНК-метилтрансферазы — группа ферментов, катализирующих метилирование нуклеотидных остатков в составе ДНК. Активность метилтрансфераз, заключающаяся в переносе метильных (CH3—) групп c кофактора(лиганда) на определенный атом азотистого основания цитозина в составе ДНК, ведет к изменению свойств ДНК, при этом изменяется активность, функции соответствующих генов, а также пространственная структура нуклеиновой кислоты (конформация).
- Запросы в Uniprot (поиск записей о метилтрансферазах человека).
- По запросу (taxonomy:"Homo sapiens (Human) [9606]" name:"dna methyltransferase") было получено 23 записи,
среди которых помимо ДНК метилтрансфераз присутствует 1 тРНК метилтрансфераза (ранее считалась ДНК метилтрансферазой, но в 2006 году было доказано, что это тРНК метилтрансфераза),
несколько белков, связывающихся с метилтрансферазой 1 (выполняют функции активации и терминации транскрипции),
похожие на них белки, связывающиеся с комплементарной ДНК (двухцепочечная ДНК, образующаяся из мРНК в результате транскрипции под действием обратной транскриптазы,
ее получают искусственно для клонирования генов эукариот в прокариотах,
а также она образуется естественным путем в результате деятельности вирусов),
также встретились члены семейства метилтрансфераз HemK (осуществляют метилирование факторов терминации трансляции у эукариот).
- Окончательный запрос (taxonomy:"Homo sapiens (Human) [9606]" name:"dna methyltransferase" NOT name:trna NOT name:cdna NOT name:hemk NOT name:associated) позволил избавиться от всех вышеперечисленных избыточных записей, получилось 9 находок.
Среди находок 5 - аннотированные (на белковом уровне), остальные неаннотированные: 2 - предсказанные, 2 получены по гомологии на уровне транскрипта.
3 аннотированные записи относятся к ДНК метилтрансферазам (DNMT1, DNMT3A, DNMT3B), катализирующим присоединение метильной группы к 5 атому азотистого основания цитозина, к этой группе, вероятно, можно отнести еще одну неаннотированную ДНК метилтрансферазу, так как имя гена всех 4 белков содержит DNMT.
- ДНК метилтрансфераза 1 метилирует цитозин в контексте CpG, предпочтительно метилирует полуметилированную ДНК. Связывается с сайтами репликации ДНК в S фазе жизненного цикла клетки для поддержания паттерна метилирования в недавно синтезированной цепи, что имеет значение для эпигенетической наследственности. Связывается с хроматином в течении фаз G2 и M для поддержания метилирования ДНК независимо от репликации. Отвечает за поддержание паттернов метилирования, закрепленных в развитии.
- ДНК метилтрансфераза 3A требуется для метилирования генома de novo и имеет важное значение для закрепления паттернов метилирования в процессе развития. Мелилирует не только CpG сайты. Играет важную роль в процессах отцовского и материнского импринтинга, требуется для метилирования самых импринтированных локусов половых клеток. Может активно подавлять транскрипцию путем набора HDAC активности.
- ДНК метилтрансфераза 3B по основным функциям схожа с ДНК метилтрансферазой 3А, а также предпочтительно метилирует ДНК в нуклеосоме и участвует в сайленсинге генов.
1 аннотированная запись соответствует метилированной-ДНК - белок-цистеин метилтрансферазе, которая участвует в клеточной защите против биологических эффектов О6-метилгуанина в ДНК. Исправлияет алкилированный гуанин путем стехиометрической передачи алкильной группы на O-6 положение остатка цистеина фермента. В ходе этой реакции фермент необратимо инактивируется. Заметим, что данный фермент осуществляет не присоединение, а отщепление метильной группы.
3 неаннотированные записи относятся к О6-метилгуанин-ДНК-метилтрансферазам, которые обладают активностью, аналогичной предыдущему белку.
Особый интерес представляет N(6)-аденин-специфичная ДНК-метилтрансфераза 2 (ДНК метилтрансферазы обычно характерны для бактерий, но в функциях сказано, что этот белок обладает потенциальной ДНК метилтрансферазной активностью, достоверных данных нет). - Ознакомиться с находками записей о ДНК метилтрансферазах и получить более полную информацию о них можно в данной таблице.
- По запросу (taxonomy:"Homo sapiens (Human) [9606]" name:"dna methyltransferase") было получено 23 записи,
среди которых помимо ДНК метилтрансфераз присутствует 1 тРНК метилтрансфераза (ранее считалась ДНК метилтрансферазой, но в 2006 году было доказано, что это тРНК метилтрансфераза),
несколько белков, связывающихся с метилтрансферазой 1 (выполняют функции активации и терминации транскрипции),
похожие на них белки, связывающиеся с комплементарной ДНК (двухцепочечная ДНК, образующаяся из мРНК в результате транскрипции под действием обратной транскриптазы,
ее получают искусственно для клонирования генов эукариот в прокариотах,
а также она образуется естественным путем в результате деятельности вирусов),
также встретились члены семейства метилтрансфераз HemK (осуществляют метилирование факторов терминации трансляции у эукариот).
- Поиск публикаций по теме в PubMed
По запросу (DNA methyltransferase[Title] AND human[Title]) было найдено 315 связанных с темой публикаций, после включения фильтра Free full text (доступный полный текст) остается 141 публикация, а вместе с фильтром Review (широкие обзоры) - 2 публикации.
Со списком публикаций по данной теме можно ознакомиться здесь.
- Поиск в PDB структур белков, относящихся к теме
По запросу (Molecule Name Search for : DNA methyltransferase and TaxonomyTree Search for Homo sapiens (human)) было получено 32 записи о структуре цепей 25 белков (содержат избыточную информацию - 5 лишних записей, так как Advanced search PDB не позволяет использовать оператор NOT).
Со списком структур можно ознакомиться в данной таблице.
- Ответы на вопросы:
- По материалам записей в Uniprot можно сделать вывод, что у млекопитающих метилируется цитозин (насчет аденина - по 6 азоту и гуанина - по 6 кислороду - трудно утверждать, так как информации об аденине найти не удалось, а что касается гуанина, фермент хоть и обладает метилтрансферазной активностью, но по гуанину осуществляет не присоединение, а отщепление -CH3). Цитозин метилируется по 5 атому и чаще всего в специфических сайтах (так называемые контексты CpG).
- Кофактором наиболее типичных ДНК метилтрансфераз (DNMT) является S-аденозил-L-метионин.
S–аденозил–L–метионин (SAMe, адеметионин) представляет собой природное вещество, производное L–метионина и аденозинтрифосфорной кислоты, которое синтезируется в печени. SAMe широко распространен во всех биологических системах организма и вовлечен в разнообразные метаболические процессы. S–аденозил–L–метионин участвует в трех наиболее важных метаболических процессах: трансметилировании, транссульфурировании и аминопропилировании. В этих реакциях он выступает либо как донор метильной группы, либо как индуктор ферментов. Значительное количество биохимических каскадов, в которых задействованы перечисленные реакции, определяет многобразие биологических эффектов SAMe.
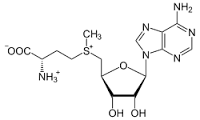
Рис. 1. S–аденозил–L–метионин.
- Стуктуры ДНК метилтрансфераз млекопитающих.
У млекопитающих обнаружены 3 активных ДНК-метилтрансферазы: DNMT1, DNMT3a и DNMT3b (ранее выделяли еще и DNMT2 (TRDMT1), но позднее была доказана тРНК метилтрансферазная активность). Также обнаружен белок, структурно схожий с семейством DNMT3, но не проявляющий метилтрансферазной активности — DNMT3L (DNMT3-like, является регуляторным фактором).- Семейство DNMT1 Основная активность этого фермента заключается в метилировании полуметилированных сайтов CpG. За предпочтение ферментом полуметилированных сайтов перед неметилированными отвечает N-концевой регуляторный участок. Считается, что ген dnmt1 образовался путём слияния гена прокариотической ДНК-метилазы с одним или двумя генами ДНК-связывающих белков. Фермент локализуется в областях репликации ДНК в течение S-фазы клеточного цикла, а после её завершения диффундирует в нуклеоплазму. Также N-концевой домен фермента DNMT1 содержит последовательность, гомологичную репрессору транскрипции HRX, с помощью которой ДНК-метилаза in vivo способна ассоциироваться с деацетилазой гистонов. Человеческий фермент DNMT1 принципиально не отличается от мышиного. Фермент DNMT1 имеет несколько изоформ: соматический DNMT1, промежуточный вариант (DNMT1b) и изоформа, характерная для ооцитов (DNMT1o).
- Семейство DNMT3 ДНК-метилтрансферазы DNMT3a и DNMT3b производят метилирование полуметилированных и неметелированных сайтов CpG с одинаковой скоростью. Гены dnmt3a и dnmt3b активно экспрессируются в недифференцированных эмбриональных стволовых клетках, тогда как в дифференцированных клетках уровень их экспрессии очень низкий. DNMT3a активнее метилирует сайты CpG, чем CpA, CpT, и CpC. DNMT3a метилирует сайты CpG намного медленнее, чем DNMT1, но быстрее, чем DNMT3b. Различия между альфа и бета формами заключаются в том, что DNMT3b отвечает за метилирование сателлитных повторов в области центромерного линкера. а мутация гена dnmt3b у человека приводит к ICF-синдрому (иммунодефицитная нестабильность центромер, аномалии лица). ICF-синдром — это редкое аутосомальное рецессивное генетическое заболевание, которое характеризуется дефектами иммунной системы и нарушением нормального строения лица. DNMT3L содержит DNA-метилтрансферазный мотив и является необходимым для эффекта материнского геномного импринтинга, оставаясь при этом каталитически неактивным (вследствие отсутствия некоторых ключевых участков, необходимых для осуществления катализа). DNMT3L экспрессируется при гаметогенезе, когда и происходит геномный импринтинг. Отсутствие DNMT3L ведет к биаллельной экспрессии генов, для которых в норме не характерна экспрессия материнского аллеля. DNMT3L взаимодействует с DNMT3a и DNMT3b в клеточном ядре. Хотя DNMT3L неспособен проводить метилирование, белок может принимать участие в репрессии транскрипции (в ассоциации с гистоновой деацетилазой).
- Биологическая роль метилирования ДНК.
- Главное значение метилирования ДНК заключается в том, что оно играет важную роль в регуляции экспрессии генов в процессе развития и дифференцировки клеток. В организмах млекопитающих метилирование ДНК обеспечивает механизм выключения больших групп генов в клетках определенного типа. Это выключение носит стабильный характер и наследуется данным типом клеток при делении (например, "выключение" одной из Х-хромосом у человека и других организмов).
- Другая его важная роль заключается в том, что метилирование - эпигенетический механизм, который коррелирует с репрессией генов, служит защитным механизмом, чтобы подавлять значительную часть генома чужеродного происхождения (т.е. реплицированные перемещающиеся элементы, вирусные последовательности и другие повторяющиеся последовательности).
- Роль метилирования у прокариот - участие в формировании системы рестрикции-модификации, благодаря которой клетки способны идентифицировать свой генетический материал и отличать его от инородных молекул, проникших в клетку тем или иным способом. Уничтожение последних позволяет поддерживать генетическую стабильность вида.
- Согласно альтернативной (эпигенетической) точке зрения, опухоль - результат неправильной работы генов (рак - болезнь регуляции генов, болезнь дифференцировки), которая может быть обусловлена метилированием ДНК. Роль метилирования ДНК в патологии наиболее явно проступает в опухолевой трансформации, когда аберрантная и стабильная инактивация генов-супрессоров приводит к тому же эффекту, что и их физическое повреждение. Это явление было названо эпимутацией, т.е. эпигенетическим эквивалентом генетической мутации. Иногда применяют термин MAGI (methylation-associated gene inactivation). Необходимо отметить, что вклад метилирования ДНК в канцерогенез не исключительно эпигенетический. В силу значительной нестабильности 5-метилцитозин способен индуцировать мутации и, как следствие, способствовать канцерогенезу хорошо известным способом - генетическим. Здесь уместно отметить, кроме того, что метилирование подавляет процесс гомологичной рекомбинации.
- Метилирование ДНК также играет роль при старении организма. Метилирование ДНК вовлечено в регуляцию активности генов. Изменения в метилировании, в частности деметилирование динуклеотидов у позвоночных, связаны с изменением уровня транскрипции, часто говорят отдельно о возрастном деметилировании. Было высказано предположение о том, что возрастное деметилирование предрасполагает клетки к опухолевой трансформации. Аберрантное метилирование CpG-островков является одной из причин, лежащих в основе изменений, наблюдаемых при старении и образовании раковых клеток.
Ссылки на источники информации: