Работа с множественными выравниваниями
В данном практикуме были выполнены задания с целью ознакомления с понятием множественного выравнивания последовательностей. В качестве объекта я использовала последовательности из файла align_07.fasta. Все задания выполнялись с помощью программы Jalview. С проектом, выполненным в данной программе, можно ознакомиться здесь.
Задание 1.
При выполнении данного задания я загрузила в программу Jalview 9 последовательностей из файла align_07.fasta, затем отсортировала их по сходству: построила дерево родства данных последовательностей (Рис. 1) (Calculate -> Calculate Tree -> Neighbour Joining Using BLOSUM62), затем отсортировала последовательности по построенному дереву (Calculate -> Sort -> By Tree Order -> Neighbour Joining Using BLOSUM62 from task_02 of Cut & Paste Input - FASTA).
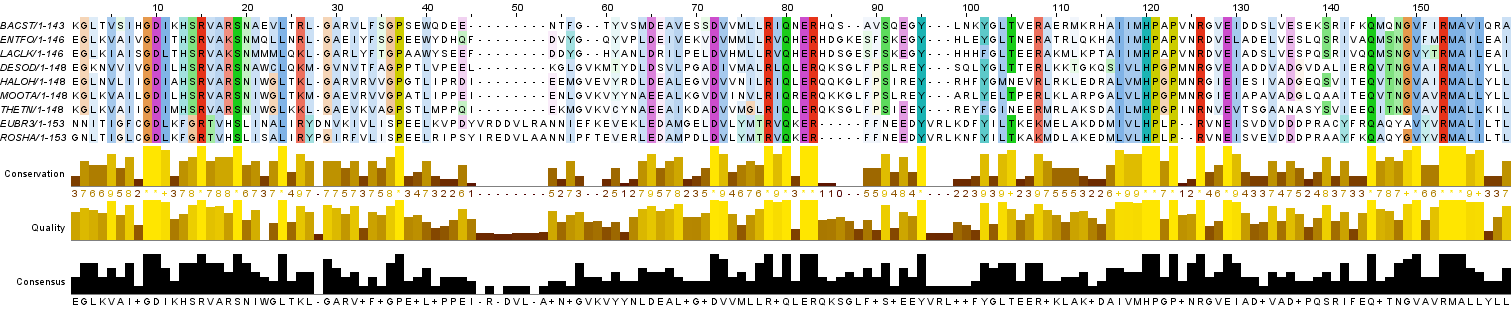
Для удобства определения консервативных и функционально консервативных позиций выравнивание было раскрашено двумя способами: ClustalX (Рис. 1a) и BLOSUM62 (Рис.1b) с порогом консервативности 30.

Задание 2.
В этом задании были найдены участки выравнивания, на которых можно ожидать гомологию аминокислотных остатков разных последовательностей. В частности, были найдены:
a) Вертикальные блоки - участки, на которых для каждой колонки можно ожидать гомологию между остатками из всех последовательностей.
В соответствии с заявленными критериями выделения вертикального блока (отсутствие гэпов, размер не менее 4 колонок, первая и последняя колонки должны быть абсолютно консервативными или функционально консервативными, не дожно быть трех не абсолютно консервативных или функционально консервативных колонок подряд) было обнаружено 9 блоков (в строке разметки Significant parts отмечены буквой "B").
Важно сказать, что для определения функционально консервативных позиций были спользованы следующие критерии свойств аминокислот: все гидрофобные (несмотря на то, что эта группа объединяет половину аминокислот), все гидрофильные (другая половина аминокислот), все ароматические, все положительно заряженные, все отрицательно заряженные, сходные по структуре доноры или акцепторы водородных связей.
По поводу использования гидрофильности и гидрофобности как критерия выделения функционально консервативных позиций: так как эти допущения увеличивают вероятность функциональной консервативности колонок, надо понимать, что если не рассматривать эти группы, то длина блоков будет значительно меньше.
Интересная особенность полученной разметки - самые обширные блоки наблюдаются в начале (22 колонки) и в конце (19 колонок) выравнивания. В середине же блоки короткие, наблюдается большое количество малоконсервативных участков, а также несколько довольно обширных гэпов.
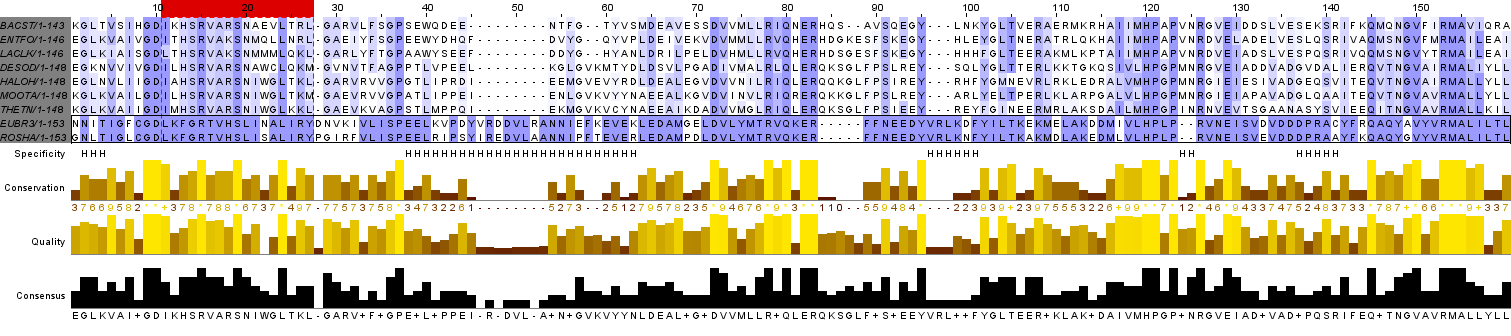
b) Кластер блоков - участок выравнивания, на котором можно ожидать гомологию между любыми двумя аминокислотами, расположенными в одной колонке.
Установленный для данного задания критерий выделения кластера таков, что два блока объединяются в кластер вместе с участком между ними, если все гэпы между этими блоками имеют длину, равную длине участка между ними.
В соответствии с этим правилом было обнаружено 4 кластера, из которых 1 представлен единственным блоком, а остальные - не менее, чем 2 блоками. На Рис. 2 кластеры блоков обозначены в строке разметки Significant parts буквой С.
Интересно, что только в одном кластере (5 - 37 колонки) блоки разделяются единственным столбцом гэпов, в остальных кластерах блоки разделяются небольшим количеством колонок без гэпов. Хочется отметить то, что в выравнивании присутствуют обширные гэпы (шириной 5 и 9 колонок), однако нельзя сказать, что колонки, окружающие эти гэпы, можно объединить в вертикальный блок, поэтому, к моему величайшему сожалению, эти гэпы не могут входить в кластеры.
c) Также на Рис. 2 в строке в строке разметки Significant parts буквой Х обозначен самый длинный участок выравнивания, не входящий ни в блоки, ни в кластеры (25 колонок).

d)На Рис. 2а черной рамкой выделена группа из двух последовательностей: EUBR3 и ROSHA, при сравнении которых с остальными последовательностями выравнивания были найдены участки, на которых остатки выбранных последовательностей, предположительно, гомологичны, а остатки остальных - скорее негомологичны им.
Причины выбора именно этих двух последовательностей:
- Они имеют одинаковую длину, отличную от длин остальных последовательностей (153 аминокислотных остатка).
- Они составляют отдельную эволюционную группу при первом же разветвлении дерева, по которому можно судить о родстве последовательностей (Рис. 1).
- Расположение гэпов таково, что они либо присутствуют во всех последовательностях внутри группы, отсутствуя за ее пределами, либо обратная ситуация.
В ходе сравнения выделенной группы с остальными последовательностями были найдены участки, на которых можно судить о гомологии между остатками выделенных в группу последовательностей, а остатки остальных последовательностей им, предположительно, негомологичны. Такие участки в строке разметки Specificity обозначены буквой H.
В позициях 12, 25, 60-63, 127, 128, 131, 132, 135, 137, 139, 143 наблюдается значимое функциональное отличие остатков внутри и вне группы, а в позициях 36, 91, 108 наблюдается такое явление, что во всех позициях вне группы остаток один, а внутри группы другой (не все эти примеры входят в состав участков, отмеченных символом Н, так как некоторые из колонок входят в состав блоков или кластеров). Именно эти примеры позволяют предположить, что на выбранных участках можно предсказать гомологию отдельно внутри группы и за ее пределами.
Задание 3.
Для первого блока (Рис. 3 - обозначен в строке разметки буквой В, 5 - 27 колонки) были определены число и процент абсолютно консервативных и функционально консервативных (согласно раскраске ClustalX), а также консервативных и функционально консервативных на 70% (возможна ситуация, когда позиция является, например, абсолютно функционально консервативной и функционально консервативной на 70% одновременно). Полученные данные представлены в Таблице 1.
Таблица 1. Число и процент различных позиций в первом блоке.
| Тип позиции | Число позиций | Процент |
| Абсолютно консервативные | 5 | 22,73% |
| Абсолютно функционально консервативные | 11 | 50% |
| Консервативные на 70% | 8 | 36,36% |
| Функционально консервативные на 70% | 5 | 22,73% |
Для самого длинного (25 колонок) участка выравнивания, не входящего в состав блоков и кластеров (Рис. 3 - обозначен в строке разметки буквой Х) были посчитаны:
- Число позиций с гэпами: 11
- Процент позиций с гэпами: 44%

Рис. 3. Изображение самого длинного участка выравнивания, не входящего в состав блоков и кластеров, а также изображение одного из кластеров.
Задание 4.
Для выполнения данного задания к выравниванию была добавлена последовательность sequence_07.fasta. Затем последовательность была вручную вписана в выравнивание с наибольшим возможным количеством значимых позиций. Результат представлен на Рис. 4.
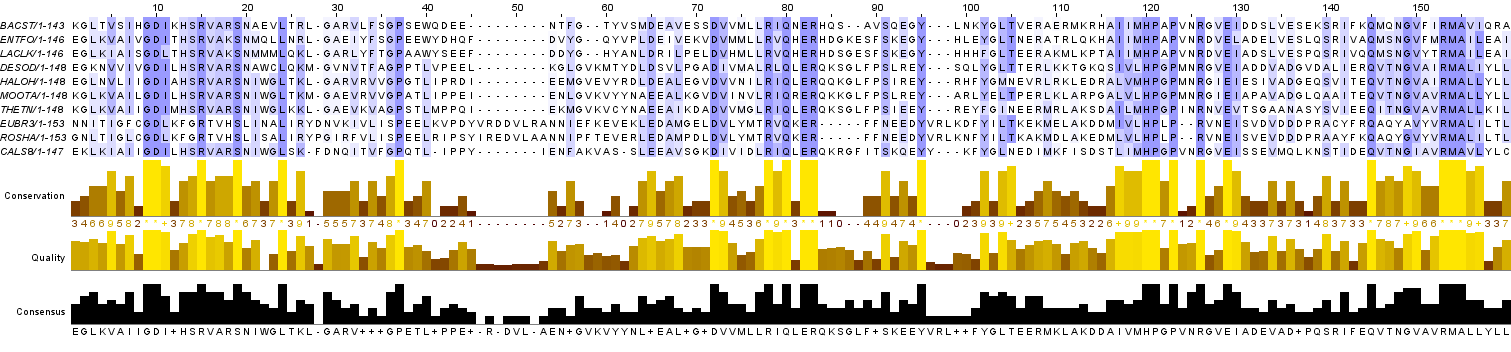
Рис. 4. Изображение выравнивания с дополнительно вписанной последовательностью из файла sequence_07.fasta.
Задание 5.
Для выполнения данного задания я добавила заведомо негомологичный участок последовательности моего белка - аспартатаминотрансферазы у организма Aquifex aeolicus VF5 (идентификатор в БД RefSeq NP_214350), так как белок слишком большой, я взяла участок длиной 152 остатка с начала последовательности.
Затем я вносила гэпы в те участки, где уже были гэпы в исходном выравнивании, с целью найти найбольшее количество совпадений с консервативными позициями в исходном выравнивании.
Удалось получить:- 2 абсолютно консервативные позиции
- 29 функционально консервативных позиций (и абсолютных, и консервативных на 70%, учитывался критерий гидрофобные/гидрофильные аминокислоты, без учета этого критерия - 17)
- 10 позиций, консервативных на 70%
- Общий процент совпадений: 26,8% ( 18,95% без учета критерия гидрофильность/гидрофобность)
Таким образом, полученный процент совпадений с конервативными позициями слишком мал, чтобы можно было судить о гомологии исходных последовательностей с добавленной последовательностью.
Результат представлен на Рис. 5.

Рис. 5. Изображение выравнивания с дополнительно вписанной последовательностью участка белка NP_214350.
Задание 6.
В этом задании было необходимо построить множественное выравнивание заведомо негомологичных последовательностей (Рис. 6) Для выполнения этого задания я взяла последовательности, с которыми работают другие студенты моего курса (идентификаторы AC в базе данных Uniprot): D9Q2Q0, B8D2N2, H6Q874, F0QTE2, F4BY60, R4T2E1. Затем я выровняла последовательность согласно инструкции.

Рис. 6. Изображение множественного выравнивания заведомо негомологичных последовательностей.
При работе с полученным выравниванием я обнаружила всего два блока, которые, за неимением конкурентов, и были признаны наилучшими (Рис. 6а и Рис. 6b).

Рис. 6a. Изображение первого наилучшего блока.

Рис. 6b. Изображение второго наилучшего блока.
Создавшаяся ситуация позволяет сделать вывод, что заведомо негомологичные последовательности имеют крайне малое число консервативных позиций, так что даже выделить блоки не всегда представляется возможным. Следовательно, судить о гомологии таких последовательностей нельзя.