Реконструкция эволюции доменной архитектуры.
Построить выравнивание представителей домена Pfam белков с разной доменной архитектурой.
Мной был выбран домен с ID CHASE. Он имеет AC PF03924.
Данный домен отвечает за связывание низкомолекулярных лигандов или небольших пептидов в рецепторе. Данный домен имеет представленные доменные архитектуры.
Для работы я выбрал две наиболее наполненные представителями доменные архитектуры: [CHASE, GGDEF] - 156 представителей и [CHASE, HisKA, HATPase_c, Response_reg] - 263 представителей.
GGDEF (PF00990) — дигуанилат-циклаза;
HisKA (PF00512) — фосфоакцепторный домен гистидин-киназ;
HATPase_c (PF02518) — домен с АТФазной активностью;
Response_reg (PF00072) — домен, воспринимающий сигналы, входящий в состав двухкомпонентной системы и располагающийся ближе к N-концу от ДНК-связывающего эффекторного домена.
Для работы был выбран тасон Proteobacteria как наиболее большой прокариотический таксон. Подтаксоны, как следует ожидать, a и b - Proteobacteria.
Excel - файл с данными для работы лежит тут.
Ссылка на проект представлена тут.
Выравнивание, полученное мной, похоже на правильное, так как домены обладают достаточной консервативностью как внутри групп, так и в целом. Все последовательности, которые были неполными/случайно попавшими были удалены. Также были удалены пустые колонки и хвосты последовательностей.
Построение филогенетического дерева домена.
Далее было произведено построение дерева. Расшифровки уже были описаны ранее, но опишу ещё раз. А - альфа-протеобактерии, В - бета-протеобактерии. 1 - первый тип доменной архитектуры (из 2 доменов). 2 - второй (из 4).
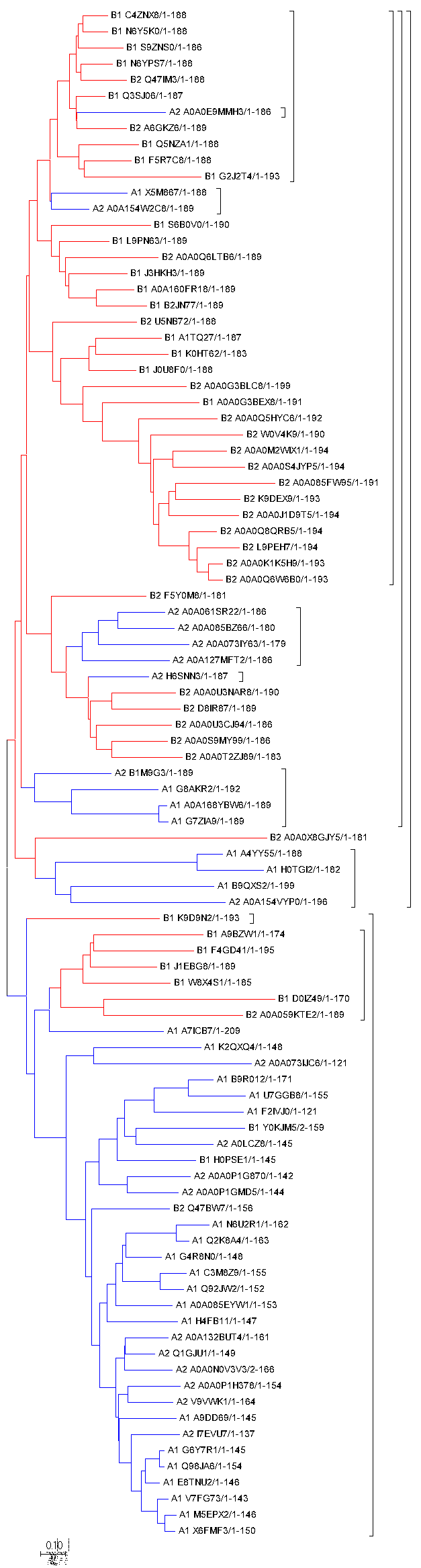
Рис.1: Дерево с окраской по таксономии. Синий - А, красный - В.
Для построения дерева использовался метод Neighbor-Joining. Скобочная структура дерева представлена здесь.
Что же мы видим на этом дереве. Во-первых хочется отметить, что у нас не получится отметить чёткие перестройки, которые отличают различные доменные архитектуры, так как, возможно, в результате ГПГ бактерии могли обмениваться фрагментами данного домена. Это привело к довольно равномерному распределению двух архитектур по дереву. В целом, можно предположить что для бета-протеобактерий родоначальниками архитектур 1 и 2 являются 1 и 3 узлы соответственно. Для альфа-протеобактерий даже такое предположение высказать трудно.
Сами таксоны достоверно различаются, однако, имеется довольно большая область перекрытия между ними, доказывающая возможность ГПГ между этими таксонами.
© Попов Алексей, 2016 г.
