Мини-обзор генома бактерии Hydrogenophilus thermoluteolus
Введение
Hydrogenophilus thermoluteolus - грамотрицательная бактерия, имеет морфотип палочки, споры не образует. Была выделена из почвы около горячего источника на полуострове Идзу, Япония, также найдена в образцах льда озера Восток, Антарктида[1]. Как видно из названия вида, колонии бактерий имеют светло-желтую окраску и их оптимальная температура роста составляет примерно 50-52°C. Оптимальный рН примерно равен 7.0 (нейтральный). Гетеротроф, использует такие органические соединения как ацетат, сукцинат, пируват и др. в качестве доноров электронов и источников углерода. Для получения азота может использовать различные ионы, кроме ионов нитритов и газообразного азота[2]. В данном мини-обзоре можно ознакомиться с описанием генома и протеома Hydrogenophilus thermoluteolus.
Систематическое положение Hydrogenophilus thermoluteolus[3]:
Домен: Bacteria
Царство: Pseudomonadati
Филум (отдел): Pseudomonadota
Класс: Hydrogenophilia
Порядок: Hydrogenophilales
Семейство: Hydrogenophilaceae
Род: Hydrogenophilus
Вид: Hydrogenophilus thermoluteolus
Материалы и методы
Данные по геному и протеому Hydrogenophilus thermoluteolus были взяты с сайта Национального Центра Биотехнологической информации (NCBI)[4]. Идентификатор генома исследуемой бактерии GCF_003574215.1_ASM357421v1. Анализ данных был проведен с использованием инструментария электронных таблиц.
Таблица с различными генами была составлена на основе таблицы геномных особенностей и функции COUNTIFS.
Для построения гистограммы длин белков шаг был выбран так, чтобы было видно примерное распределение длин белков, а «хвост» гистограммы обрезан на значении, данные после которого очень малы и почти не несут информации.
Способность к хемолитоавтотрофии была проанализирована с помощью фильтра названий генов и их отбора, с учетом знания названий ферментов, исследуемых циклов.
Результаты
1. Типы различных генов и их количество
Геном исследуемой бактерии Hydrogenophilus thermoluteolus представлен одной хромосомой и одной плазмидой. Всего было найдено 5 типов генов (см. сопроводительные материалы, S1). CDS – coding sequences, ncRNA – некодирующие РНК, tRNA – транспортные РНК, rRNA – рибосомальные РНК, tmRNA – транспортно-матричная РНК, нужна для высвобождения рибосом, «застрявших» на проблемных участках мРНК[5]. Все гены РНК располагаются на хромосоме, как и большинство CDS. В таблице 1 представлено количество каждой из них на хромосоме и плазмиде.
Можно отметить, что все гены РНК содержатся исключительно на хромосоме, так как плазмида несет только добавочную информацию, которая не является жизненно необходимой. Количество CDS на плазмиде так же очень мало, вероятно, это гены устойчивости к чему-либо, которые ей помогают приспосабливаться.
Заметим, что число генов рРНК превышает ожидаемое, возможно, это произошло за счет дублирования. Количество генов тРНК наоборот меньше ожидаемого (49 вместо 61 предположительных). Причины этого могут быть различны. Один из вариантов – избегание бактерией определенных кодонов, в результате чего будут отсутствовать некоторые тРНК. Также возможно неполное соответствие кодонов и антикодонов (например, в некоторых случаях могут совпадать только два нуклеотида из трех). При дальнейшем анализе могут быть сделаны довольно интересные наблюдения.
| Тип репликона | CDS | ncRNA | tRNA | rRNA | tmRNA |
|---|---|---|---|---|---|
| chromosome | 2104 | 3 | 49 | 9 | 1 |
| plasmid | 76 | 0 | 0 | 0 | 0 |
2. Анализ длин белков, закодированных в геноме Hydrogenophilus thermoluteolus
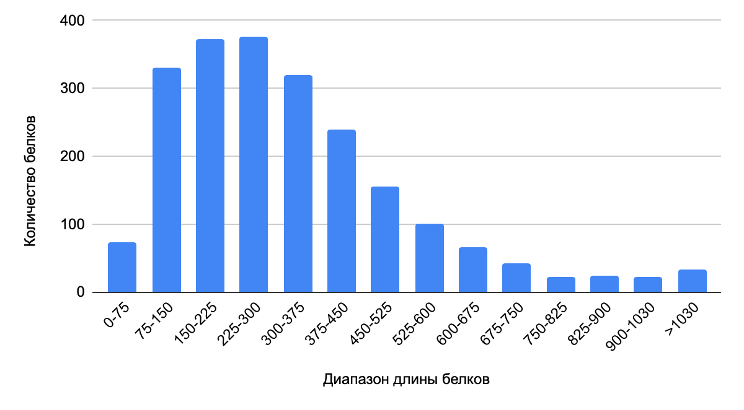
Всего было проанализировано 2181 возможных продуктов трансляции CDS (см. сопроводительные материалы, S2). Ниже представлена диаграмма, отображающая распределение длин белков.
Как видно из рисунка, большинство белков Hydrogenophilus thermoluteolus имеют длину 150-300 аминокислот, а медианное значение составляет 286 аминокислот.
3. Анализ GC%-состава генома
На основании данных таблицы особенностей генома было посчитано среднее процентное содержание пар GC в составе CDS (см. сопроводительные материалы, S3). В хромосоме оно составило 61,62%, а в плазмиде – 58,41%. Из этого можно сделать вывод, что бактерия обитает при высоких температурах (термофильная), так как GC пары являются более крепкими за счет трех водородных связей, в отличие от AT пар, где водородных связей по две. Это согласуется с найденной о бактерии информацией.
4. Способность к хемолитоавтотрофии
В ходе работы была поставлена задача изучить способность Hydrogenophilus thermoluteolus к хемолитоавтотрофии (см. сопроводительные материалы, S4). По найденным данным род Hydrogenophilus описывается как хемолитоавтотрофные бактерии, то есть они имеют способность использовать в качестве источника углерода СО2 из воздуха и неорганические вещества в качестве донора электронов[1]. Однако Hydrogenophilus thermoluteolus считается гетеротрофным организмов.
С помощью таблицы особенностей генома был проведен анализ продуктов трансляции и по названиям выявлены различные ферменты. Так на минус-цепи обнаруживаются последовательности, кодирующие ферменты цикла Кальвина (нужен для фиксации СО2) такие как рибулозо-1,5-бисфосфаткарбоксилаза, глицеральдегид-3-фостфат дегидрогеназа и другие. Кроме того, были найдены CDS различных гидролаз, которые позволяют использовать водород как источник электронов. Таким образом, Hydrogenophilus thermoluteolus является примером гетеротрофной бактерии, которая при необходимости может становиться хемолитоавтотрофной (факультативный хемолитоавтотроф).
Сопроводительные материалы
Для ознакомления со всеми данными и результатами можно перейти по следующим ссылкам:
- Лист «Типы кодирующих последовательностей» (S1)
- Лист «prot_lengths_hist» (S2)
- Лист «GC%» (S3)
- Лист «Хемолитоавтотрофия» (S4)
Список литературы
- Sergey A. Bulat; Irina A. Alekhina; Michel Blot; Jean-Robert Petit; Martine de Angelis; Dietmar Wagenbach; Vladimir Ya. Lipenkov; Lada P. Vasilyeva; Dominika M. Wloch; Dominique Raynaud; Valery V. Lukin, DNA signature of thermophilic bacteria from the aged accretion ice of Lake Vostok, Antarctica: implications for searching for life in extreme icy environments, International Journal of Astrobiology, 2004, vol. 3, no. 1.
- Nobuhiro R. Hayashi, Tatsuya Ishida, Akira Yokota, Tohru Kodama and Yasuo Igarashi, Hydrogenophilus thermoluteolus gen. nov., sp. nov., a thermophilic, facultatively chemolithoautotrophic, hydrogen-oxidizing bacterium, International Journal of Systematic and Evolutionary microbiology, 1999, vol. 49, no. 2