Поиск сигналов I
Задание выполнялось Карпухиной Анной и Буяновой Мишель.Определение биологической роли TF в бактерии
Работа проводилась с бактерией Bacillus pumilus SAFR-032. Файл LiaR.fasta с участками связывания транскрипционного фактора LiaR доступен по ссылке.
Поиск мотива связывания производился при помощи сервиса MEME. Параметры находки приведены в Таблице 1.
Таблица 1. Мотив, найденный MEME
| Мотив | LOGO | PWM | E-value |
ATMCGMCTYNRGTCKKA |  | PWM.txt | 5.5e-036 |
Далее с использованием сервиса tomtom проводился поиск похожего мотива по БД RegTransBase. Параметры лучшей находки (со схожей PWM) приведены в Таблице 2.
Таблица 2. Находка tomtom
| Название ТФ | Длина мотива находки | Alignment | PWM | E-value |
| NarP | 16 | 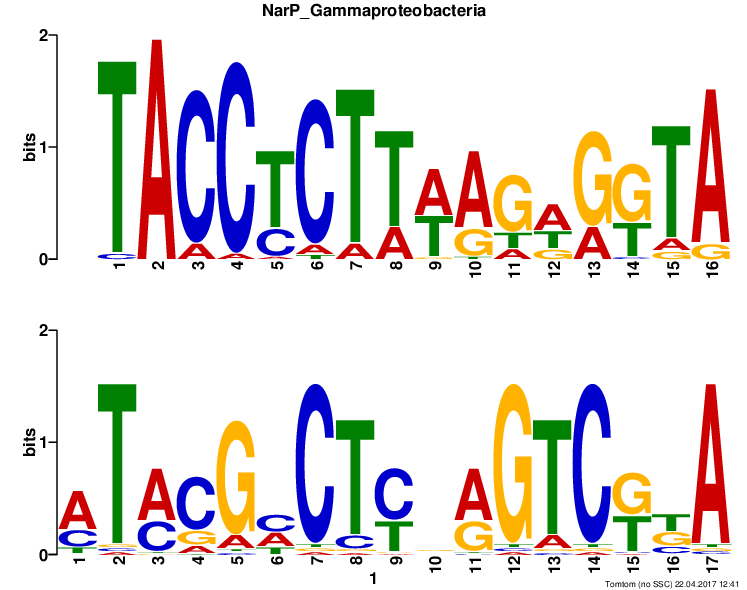 | tomtom_PWM.txt | 5.58e-02 |
Выравнивание, приведенное в ячейке Alignment, позволяет наглядно сравнить мотив-находку с мотивом, найденным MEME. Так, например, мотив из tomtom короче исходного на 1 нуклеотид, и его первый нуклеотид выровнен с позицией 2 исходного. Также стоит указать,
что ориентация найденного мотива — Reverse Complement.
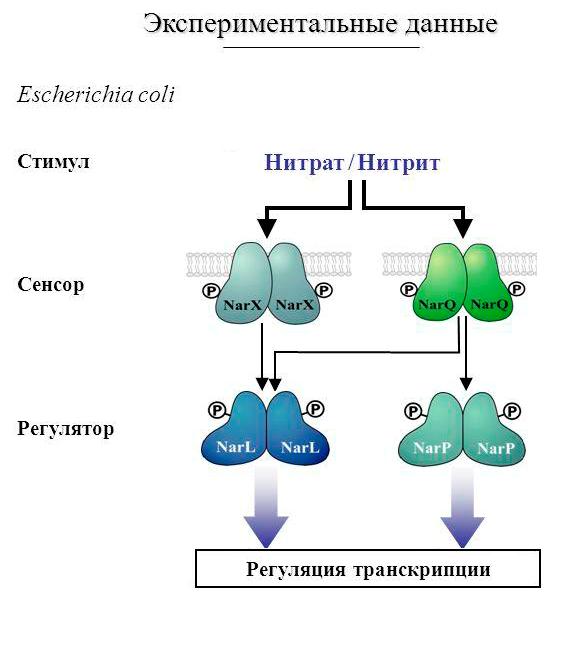
Интересно отметить, что NarP в бактериях отвечает за регуляцию нитрат-нитритного дыхания (см. Рис. 1) и входит в состав двойной системы NarP/NarQ, а сайт его связывания представляет собой палиндром.[1]
Рис. 1. Роль NarP в дыхании гаммапротеобактерий[1]
Исходно найденный мотив был передан сервису FIMO. В качестве БД, по которой проводилось сканирование на поиск схожих мотивов, была выбрана Upstream Prokaryotic для исследуемой нами бактерии Bacillus pumilus SAFR 032.
Все находки FIMO в текстовом формате лежат в fimo.txt. Информация же об исключительно координатах и p-value находок представлена в [xlsx]-файле fimo.xlsx
и, для наглядности, в Таблице 3.
Таблица 3. Параметры находок FIMO
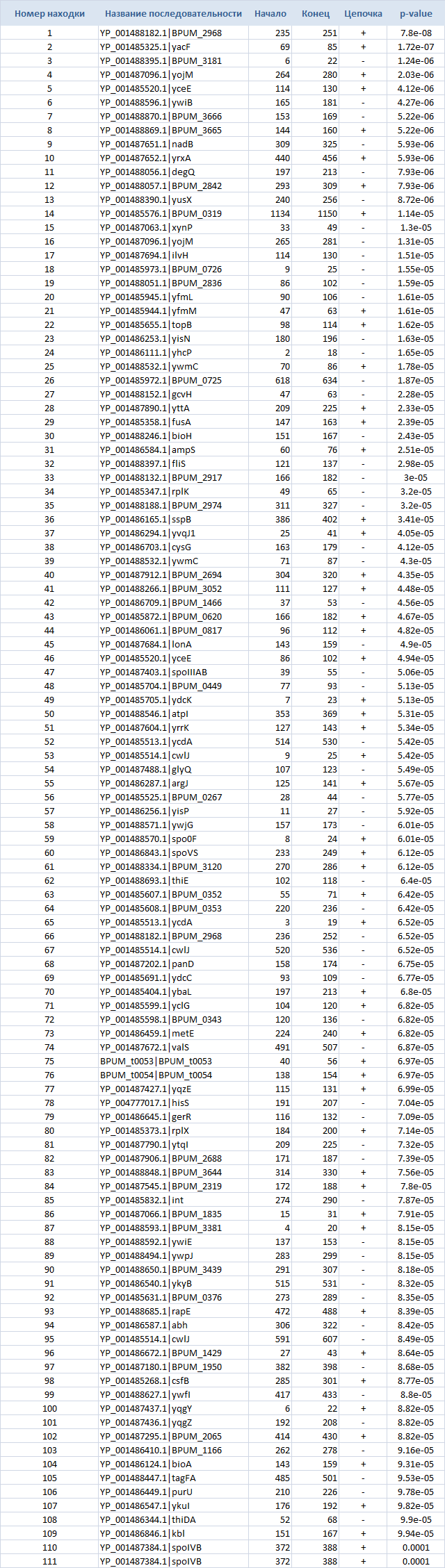
Объяснение тому, что поиск проводился только по upstream regions, довольно простое и заключается в том, что связывание факторов транскрипции происходит по большей части именно в участке, предшествующему гену, т.е. в upstream, для того чтобы иметь влияние на экспрессию гена (подавлять ее или способствовать ей, выполняя тем самым свою роль).
Выбор БД облегчил задание по поиску генов, на экспрессию которых может влиять рассматриваемый ТФ. Все находки FIMO, помимо двух, обладали непозволительно высоким q-value (~1). Именно поэтому такие хиты были отфильтрованы,
и в итоге найденные гены — это BPUM_2968 и yacF (их q-value, честно говоря, тоже не позволяет говорить о достоверности: 0.185, 0.203).
Программой featcopy были извлечены координаты генов в геноме Bacillus pumilus и по ним восстановлены координаты upstream regions: complement(22953414..2953749) и 66535..66713. Последовательности данных участков использовались в Задании 2.
Анализ с помощью БД String[2] позволил установить, что гены, предположительно регулируемые нашим ТФ, скорее всего, не входят в один метаболический путь. Что касается функций этих генов, то об их схожести также сложно что-либо сказать. Ген yacF кодирует белок тРНК-дигидроуридин синтазу, катализирующую синтез 5,6-дигидроуридина (D) - модифицированного основания, встречающегося в D-петле большинства тРНК. Ген BPUM_2968 кодирует неохарактеризованный пока белок, о котором известно лишь то, что он является трансмембранным.
Пересечение сайтов связывания транскрипционного фактора LiaR c сайтами узнавания бактериальных ДНК-метилтрансфераз
Метилирование оснований в ДНК имеет место во всех пяти царствах живых существ, включая бактерий. Наиболее типичное метилированное основание, встречающееся повсеместно - C5-Метил-цитозин (m5C). Для бактерий, помимо этого, характерно наличие в ДНК N6 -метил-аденина (m6A) (есть у бактерий и низших эукариот) и N4-метил-цитозина (m4C) (исключительно бактериальный). Метильные группы m6A, m4C and m5C выступают из большой бороздки двойной спирали, которая является основным местом расположения сайтов связывания различных ДНК-связывающих белков. Поэтому метилирование A-C мотивов может в занчительной степени влиять на ДНК-белковые взаимодействия.
Формирование m6A, m4C и m5C катилизируется ДНК-метилтрансферазами, имеющимим собственные сайты узнавания. Подавляющая часть описанных к настоящему времени ДНК-метилтрансфераз являются участниками систем рестрикции-модификации, каждая из которых в типичном случае состоит из эндонуклеазы рестрикции и соответсвующей метилтрансферазы, "защищающей" необходимые участки от разрезания этой эндонуклеазой. Тем не менее существуют и одиночные метилтрансферазы, которые, вероятно, произшли в следствие утраты соответсвующих эндонуклеаз (наиболее известный пример - Dam-метилаза гамма-протеобактерий).
Если сайт связывания ДНК-метилазы пересекается с промоторным или регуляторным участком, метилирование может оказывать влияние на связывание транскрипционных факторов и, соответственно, модулировать транскрипцию. Метилирование участков, расположенных в относительной близости от сайта связвания транскрипционного фактора также может иметь подобный эффект[3].
С помощью программы fuzznuc пакета EMBOSS был проведен поиск сайтов метилирования, пересекающихся с найденными FIMO мотивами.
Для этого был взят файл с известными последовательностями сайтов метилирования (MT-sites) и файлы, содержащие два лучших мотива и их окрестности в 50 нуклеотидов с каждой стороны (чтобы не упустить пересечения с сайтами, превышающими длину мотива) (mot1.fasta, mot2.fasta). Использованная команда:
fuzznuc -sequence mot1.fasta -pattern @MT-sites -outfile fuzznuc1.out
В результате работы программы нашлось достаточно много участков пересечения для каждого из мотивов (505 и 370), однако примерно треть находок в обоих случаях представляли собой 1-2 буквенные последовательности, которые, на наш взгляд, не несут особого смысла. Если исключить их из числа значимых находок, то фактическое число потенциальных участков пересечения становится гораздо меньше (285 и 225). Обобщение полученных данных представлено в Таблице 4.
| Ген | Координаты мотива с окрестностью в 50 нуклеотидов | Общая длина участка | Количество находок | Выходной файл |
| BPUM_2968 | complement(2953364..2953799) | 435 | 505 | fuzznuc1.out |
| yacF | 66485..66763 | 278 | 370 | fuzznuc2.out |
В целом пересечений последовательностей наших мотивов и сайтов связывания метилтрансфераз достаточно много. Тем не менее говорить однозначно о том, что метилирование (если оно вообще действительно происходит в этих участках) как-то влияет на транскрипцию изучаемых генов, достаточно сложно. Многие пересечения короткие и/или содержат протяженные участки, в которых нуклеотиды могут варьироваться (IUPAC Ambiguity Codes), поэтому они могли найтись случайно. Некоторые пересечения могли попасть в окрестность и не затронуть сам мотив.
Самые длинные пересечения, не включающие N-позиций, имеют длину 6 нуклеотидов, более длинные пересечения, как правило, имеют посередине протяженный N-участок (например CCANNNNNNTTC - CCATTCATTTTC). Достаточно часто встречаются повторяющиеся пересечения. Интересно, что по координатам они часто идут друг за другом.
Чтобы проверить есть ли в геноме нашей бактерии ДНК метилтрансферазы с той же специфичностью, что у тех, для которых были найдены пересечения с мотивами, геном Bacillus pumilus SAFR-032 был найден в базе данных REBASE и была получена информация о системе рестрикции-модификации данной бактерии (Рис. 2). Имеющиеся метилтрансферазы обозначены фиолетовым, обе они являются метилтрансферазами второго типа.
Рис. 2. Выдача REBASE
Первая из них — M.Bpu32ORF561P (Рис. 3) является цитозин-5 ДНК метилтрансферазой, участок ее узнавания неизвестен.
Риc. 3. M.Bpu32ORF561P
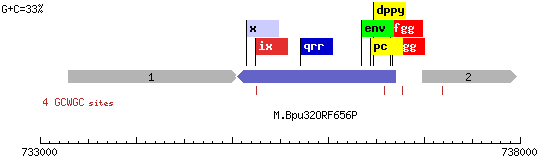
Вторая же, — M.Bpu32ORF656P (Рис. 4), — является цитозин-5 ДНК метилтрансферазой с предполагаемым участком узнавания GCWGC (W=A/C).
Рис. 4. M.Bpu32ORF656P
Для первого мотива данный сайт в найденных участках не присутствовал, однако были похожие сайты на 1 нуклеотид длиннее - GCATGC и GCAAGC. Для второго мотива самое похожее, что было найдено - GCATC, то есть mismatch в один нуклеотид. Так как точных совпадений найдено не было, вероятно, что метилирование не имеет серьезного влияния на связывание ТФ в нашем случае.
[1] А.В. Герасимова, Д.А. Равчеев. Компьютерное предсказание потенциальных участков связывания FNR и ArcA // Международная научная конференция студентов, аспирантов и молодых учёных «Ломоносов-2004», стр. 10, Москва, Россия, Апрель 2004.
[2] STRING
[3] DNA methylation in bacteria: from the methyl group to the methylome, Sánchez-Romero MA, Cota I, Casadesús J.