Анализ генома и протеома Pseudomonas alkylphenoliсa
Е.С. Песковацкова1*
1Факультет биоинженерии и биоинформатики, Московский государственный университет им. Ломоносова, Россия, Москва
СОДЕРЖАНИЕ
Был проведен анализ геномной последовательности и анализ кодирующих последовательностей Pseudomonas alkylphenoliсa. В результате чего стали известные стандартные данные о геноме Pseudomonas alkylphenoliсa: число и названия ДНК, составляющих геном; их длина и GC-состав; координата начала репликации (oriC) и координата терминации реплиции (ter). Были найдены частоты использования стоп-кодонов и отмечены свойства кодирующих последовательностей с редко-встречающимися стоп-кодонами, а также проанализированы последовательности, в которых стоп-кодон не занимает концевое положение. Работая с протеомом бактерии, были найдены длины белков, общее количество использований каждой аминокислоты в протеоме; были найдены частоты использования кодонов, кодирующих одну и ту же аминокислоту. Полученные результаты согласуются с имеющейся на данный момент информацией о геноме и протеоме Pseudomonas alkylphenoliсa.
1 ВВЕДЕНИЕ
Pseudomonas alkylphenoliсa - штамм анаэробных бактерий, представители которого впервые были найдены в почве, содержащей с1-с5 алкилфенолы. Несмотря на наличие названия Pseudomonas alkylphenolica, данный штамм до сих пор не имел четкой таксономической характеристики [1]. Но всё же проведенные исследования позволяют классифицировать эти бактерии как штамм нового вида - KL28T [1].
Pseudomonas alkylphenoliсa имеют один полярный жгутик, за счет чего достигают достигают длины в 3.1–3.5 μm . Их клетки дают отрицательный результат на каталазный и оксидазный тесты; аэробные , не ферментативные [1] . Существование в агрессивной среде - результат образования разных клеточных форм в зависимости от внешних условий. В частности на среде, содержащей пара алкил-фенолы, они формируют воздушные структуры за счет lap - катаболического генного кластера[2].
Такая устойчивость к алкилфенольной среде , позволила культивировать данный штамм с использованием пара-крезола и заметить его потенциал как в качестве объекта исследований индивидуального развития бактерий, так и как в качестве биофильтра [1].
2 MЕТОДЫ
В ходе анализа шла работа с файлами, содержащими информацию о геноме, кодирующих последовательностях (cds) или протеоме Pseudomonas alkylphenoliсa. В основном использовались методы программирования на Python, возможности сервиса Google-sheets (для создания электронных таблиц и гистограмм).
В ходе работы с геномом Pseudomonas alkylphenoliсa:
Стало известно число и название днк, составляющих геном благодаря файлу с геномной последовательностью (файл 1).
Длина днк и гц-состав были подсчитаны - благодаря написанным программам (pr.1 и pr.2 соответственно )
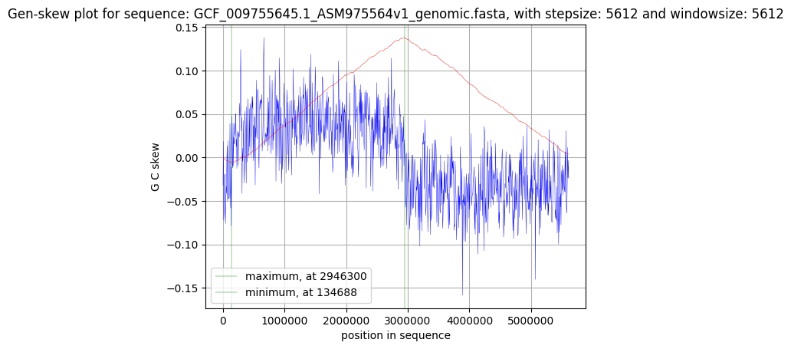
Координата начала репликации (oriC) и координата терминации реплиции (ter) (см. гр. 1) были найдены с помощью онлайн-сервиса для вычисления GC-skew на основе данных файла с геномной последовательностью (файл 1)
Для определения частоты использования определенных старт-кодонов (таб 1.) была написана программа (pr. 3). Также использовался файл с cds (файл 2)
Была написана программа (pr. 4) для подсчета встречаемости стоп-кодонов в cds – результат отражен в таб.2. Также, с помощью (pr. 5) найдены кодирующие последовательности, в которых стоп-кодон не занимал концевое положение.
Действия, выполненный в ходе работы с протеомом Pseudomonas alkylphenoliсa:
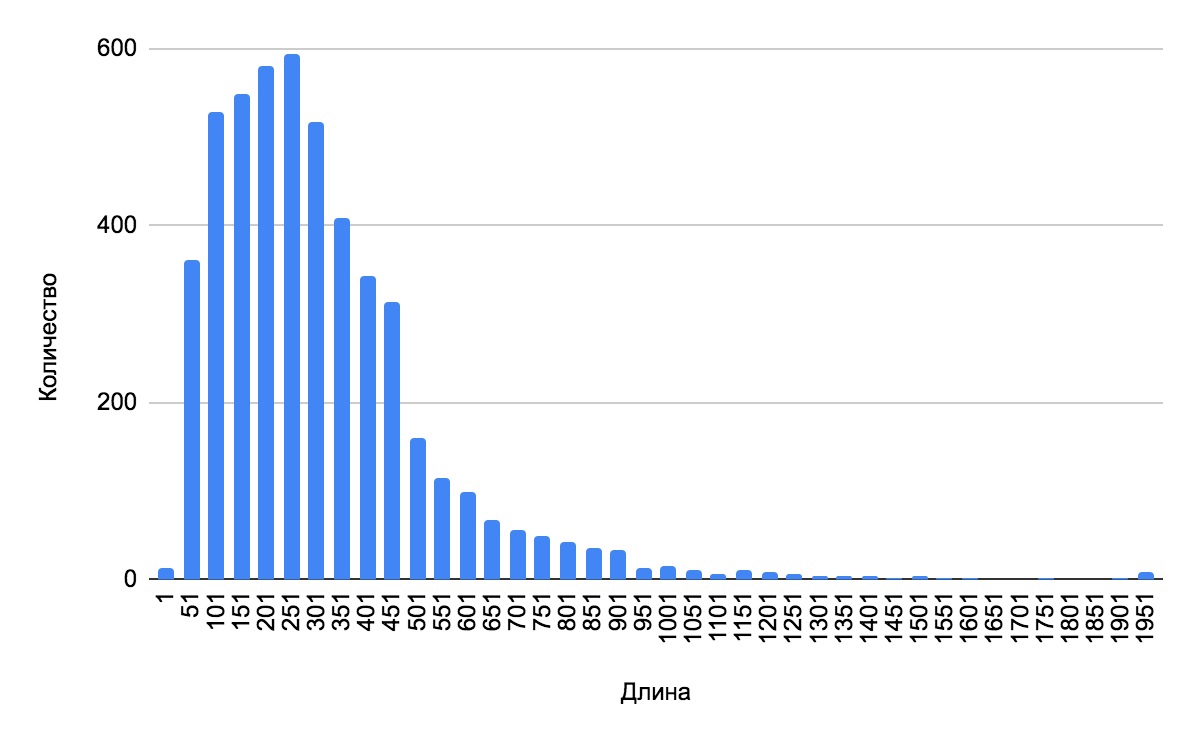
с помощью функций Google-Sheets, на основе таблицы особенностей генома (таб.0) была построена гистограмма (гист. 1), отражающая распределения длин белков бактерии.
На основе cds с помощью написанной программы (pr. 6) была также составлена таблица использования кодонов. кодирующих одну и ту же аминокислоту (далее - АК))- (таб.3) и (таб. 4)
По данным файла с протеомом бактерии (файл 3) с помощью написанной программы (pr.7) было выяснено количественное соотношение АК в протеоме Pseudomonas alkylphenoliсa.
3 РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
3.1 Стандартные данные о геноме:
Геном Pseudomonas alkylphenoliсa состоит из одной хромосомы - Neo, которая содержит в себе 1 молекулу днк.
Длина единственной хромосомы - 5 612 010 bp (base-pairs — пар оснований). Среднее GC содержание - 0.6118.
Полученные данные соответствуют действительности [3].
GC содержание Pseudomonas alkylphenoliсa сопоставимо с GC содержанием Pseudomonas fluorescens SBW25 [4].
3.2 Координата начала репликации ( oriC) и координату терминации репликации (ter.)
график. 1. GC-skew Pseudomonas alkylphenoliсa
Так как в некоторых геномах прокариот на лидирующей цепи гуанина больше чем цитозина, а на запаздывающей цепи больше цитозина чем гуанина, то можно воспользоваться методом GC-skew, основанном на этом наблюдении, для того, чтобы определить координату начала репликации (oriC) и координату конца репликации (ter).
Формула для расчета cumulative gc-skew:
GC_skew = (G_cnt - C_cnt)/(G_cnt + C_cnt)
формула 1.
где G_cnt и C_cnt - соответственно количество гуанинов и цитозинов в рассматриваемом “окне” – произвольно выбранном диапазоне нуклеотидов. Окно продвигается по последовательности с произвольно выбранным “шагом”.
Таким образом координата oriC соответствует минимальному значению cumulative gc-skew – 134688, координата ter. соответствует максимальному значению cumulative gc-skew – 2946300.
3.3. Частота использования определенных старт кодонов.
старт-кодоны
краткое описание соответствующей кодирующей последовательности
ACG 1
псевдоген
ATA 10
1. белок из супер-семейства белков посредников. Участвует в синтезе гемолизина 2. 16s р-РНК метилтрансфераза 3.вставочный мембранный белок. 4.белок трансмембранного транспорта. 5. Белок, участвующий в разрушении пиримидиновых оснований. 6,7 внеклеточный белок, связывающий растворенные вещества. 8.Катализирует НАД-зависимое восстановление полуальдегида сукцинилглутамата в сукцинилглутамат. 9. белок супер-семейства метал-зависимых и метал не зависимых фосфогидролаз[5]. 10. Пеницилл-зависимый белок, участвует в формировании клеточной стенки, синтез пептидогликанов из промежуточных продуктов липидов.
Видно, что старт кодоны с наименьшей встречаемостью находятся в кодирующих последовательностях псевдогенов. Оставшиеся кодоны можно трактовать как синонимичные замены, которые не влияют на реализацию информации, или как способ регулирования процесса трансляции.
как было сказано ранее, среднее GС- содержание Pseudomonas alkylphenoliсa – 0.6118. В то же время TGA – самый часто-используемый стоп-кодон в геноме бактерии, и , согласно имеющимся данным, прослеживается прямая зависимость между высоким средним GC - содержанием и количеством TGA стоп-кодонов[6].
Последовательности, в которых стоп-кодон не занимал конечного положения.
Также были найдены 17 кодирующих последовательностей, в которых стоп-кодон не занимал конечного положения. Среди них лишь одна не являлась последовательностью псевдогена. Она кодирует N-субъединицу формиатдегидрогеназы, (функция которой состоит в том, чтобы катализировать окисление формиата до СО2). В ней TGA считывается не как стоп-кодон, а как кодон, отвечающий за синтез селеноцистеина, благодаря специфичной последовательности после него.[7]
3.5. Распределение длин белков.
гист. 1.распределение длин белков.
В среднем белки Pseudomonas alkylphenoliсa имеют длину в 251 аминокислотный остаток, что вполне типично для прокариот[8]. Что касается самых длинных белков, чья длина превышает 501 аминокислотный остаток, то большинство из них будут иметь отношение к клеточным процессам и клеточному метаболизму [8]. И , благодаря таблице особенностей генома Pseudomonas alkylphenoliсa, можно убедиться в этом.
Таким образом, распределение длин белков организма может дать примерную оценку количеству белков выполняющих разные функции.
3.6. Частота использования кодонов, кодирующих одну и ту же аминокислоту.
а/к
кодоны
кол-во
сумма
Ala, A
GCT
36 171
min
36 171
194 541
GCC
59 877
max
59 877
GCA
43 221
GCG
55 272
Arg, R
CGT
21 572
min
18 270
201 903
CGC
52 658
max
52 658
CGA
34 355
CGG
43 904
AGA
18 270
AGG
31 144
Asn, N
AAT
9 355
min
9 355
33 849
AAC
24 494
max
24 494
Asp, D
GAT
19 266
min
19 266
41 145
GAC
21 879
max
21 879
Cys, C
TGT
22 030
min
22 030
78 349
TGC
56 319
max
56 319
Gln, Q
CAA
23 459
min
23 459
53 661
CAG
30 202
max
30 202
Glu, E
GAA
18 951
min
15 350
34 301
GAG
15 350
max
18 951
Gly, G
GGT
25 024
min
16 439
111 954
GGC
48 778
max
48 778
GGA
16 439
GGG
21 713
His, H
CAT
17 197
min
17 197
38 737
CAC
21 540
max
21 540
Ile, I
ATT
10 505
min
5 427
41 638
ATC
25 706
max
25 706
ATA
5 427
Leu, L
CTT
14 242
min
4 976
109 732
CTC
14 548
max
46 852
CTA
8 299
CTG
46 852
TTA
4 976
TTG
20 815
Lys, K
AAA
16 090
min
16 090
45 360
AAG
29 270
max
29 270
Met, M
ATG
26 550
26550
Phe, F
TTT
10 579
min
10 579
31 324
TTC
20 745
max
20 745
Pro, P
CCT
32 828
min
31 620
157 100
CCC
31 620
max
55 177
CCA
37 475
CCG
55 177
Ser, S
TCT
15 525
min
14 541
173 361
TCC
20 554
max
48 328
TCA
33 626
TCG
48 328
AGT
14 541
AGC
40 787
Thr, T
ACT
16 754
min
16 754
110 942
ACC
40 196
max
40 196
ACA
24 030
ACG
29 962
Trp, W
TGG
56 485
56485
Tyr, Y
TAT
7 159
min
7 159
20 961
TAC
13 802
max
13 802
Val, V
GTT
18 490
min
11 097
83 111
GTC
22 248
max
31 276
GTA
11 097
GTG
31 276
таб. 3. частота использования кодонов, кодирующих 1 аминокислоту.
Из таблицы видно, что самый часто-используемый код-
он – GCC (кодирует аланин) – встречается 59877 раза, а самый редко-используемый – TTA (кодирует лейцин ) — 4976 раз. Стоит отметить что, такое распределение кодонов можно связать с высоким средним GC-содержанием [9].
И, если говорить про количество кодонов имеющих вид “*GC” или “*CG” ( где ‘*’ - это любой из нуклеотидов), то их в геноме Pseudomonas alkylphenoliсa значительно больше, чем кодонов оканчивающихся на “А” и “T”, и это также можно связать с относительно высоким средним GC содержанием:
*GC
сумма
*GC
сумма
*GC+*CG
GCC
59 877
313 691
GCG
55272
285301
598 992
GCG
55 272
CGC
52658
CGC
52 658
CGG
43904
TGC
56 319
CCG
55177
GGC
48 778
TCG
48328
AGC
40 787
ACG
29962
*AT
сумма
*TA
сумма
*AT+*TA
AAT
9 355
58 404
ATA
5427
50760
109 164
GAT
19 266
CTA
8299
CAT
17 197
TTA
4976
ATA
5 427
TAT
7159
TAT
7 159
TAC
13802
GTA
11097
таб. 5. Количественное соотношение аминокислот в протеоме.
В протеоме мало аминокиcлот с ароматическим радикалом (F, Y, W) . Это может быть одной из причин, по которой Pseudomonas alkylphenoliсa выживает на алкилфенольной среде. А малое количество цистеина связано с тем, что свободный цистеин может быть токсичен для клеток прокариот и его синтез находится под жестким контролем [10].
Таким образом, рассматривая количественное соотношение аминокислот в протеоме, можно сделать предположения о характеристиках организма.
3.7. Распределение длин белков.
А/К
кол-во
A, Ala
185 381
R, Arg
106 749
N, Asn
51 164
D, Asp
88 261
C, Cys
16 961
Q, Gln
81 253
E, Glu
94 520
G,Gly
132 416
H, His
37 584
I, Ile
78 673
L, Leu
200 338
K, Lys
56 417
M, Met
37 771
F, Phe
60 211
P, Pro
79 777
S, Ser
98 209
T, Thr
78 737
W, Trp
24 229
Y, Tyr
43 072
V, Val
118 654
max (лейцин)
200 338
min (цистеин)
16 961
таб. 5.Количественное соотношение аминокислот в протеоме
В протеоме мало аминокиcлот с ароматическим радикалом (F, Y, W) . Это может быть одной из причин, по которой Pseudomonas alkylphenoliсa выживает на алкилфенольной среде. А малое количество цистеина связано с тем, что свободный цистеин может быть токсичен для клеток прокариот и его синтез находится под жестким контролем [10].
Таким образом, рассматривая количественное соотношение аминокислот в протеоме, можно сделать предположения о характеристиках организма.
ЗАКЛЮЧЕНИЕ
В обзоре были рассмотрены некоторые свойства генома и протеома Pseudomonas alkylphenoliсa. Говоря про геном, стоит отметить, что большинство рассмотренных свойств зависели от GC-содержания и принципа случайного распределения кодонов.
Сравнение длины белков, вычисление встречаемости кодонов, кодирующих 1 аминокислоту, и подсчет количества соответствующих аминокислот дали краткие сведения о протеоме Pseudomonas alkylphenoliсa.
Полученные результаты соотносятся с имеющейся на данный момент информацией об особенностях генома и протеома Pseudomonas alkylphenoliсa.
СПИСОК ЛИТЕРАТУРЫ
Mulet M, Sánchez D, Lalucat J, Lee K, García-Valdés E. Pseudomonas alkylphenolica sp. nov., a bacterial species able to form special aerial structures when grown on p-cresol. Int J Syst Evol Microbiol. 2015;65(11):4013-4018. doi:10.1099/ijsem.0.000529
Lee K, Lim EJ, Kim KS, Huang SL, Veeranagouda Y, Rehm BH. An alginate-like exopolysaccharide biosynthesis gene cluster involved in biofilm aerial structure formation by Pseudomonas alkylphenolia. Appl Microbiol Biotechnol. 2014;98(9):4137-4148. doi:10.1007/s00253-014-5529-6
Brocchieri L, Karlin S. Protein length in eukaryotic and prokaryotic proteomes. Nucleic Acids Res. 2005;33(10):3390-3400. Published 2005 Jun 10. doi:10.1093/nar/gki615
Lightfield J, Fram NR, Ely B. Across bacterial phyla, distantly-related genomes with similar genomic GC content have similar patterns of amino acid usage. PLoS One. 2011;6(3):e17677. Published 2011 Mar 10. doi:10.1371/journal.pone.0017677
Alexander T Ho, Laurence D Hurst, Variation in Release Factor Abundance Is Not Needed to Explain Trends in Bacterial Stop Codon Usage, Molecular Biology and Evolution, Volume 39, Issue 1, January 2022, msab326, https://doi.org/10.1093/molbev/msab326
Yoshizawa, S., & Böck, A. (2009). The many levels of control on bacterial selenoprotein synthesis. Biochimica et Biophysica Acta (BBA)-General Subjects, 1790(11), 1404-1414.
Luciano Brocchieri, Samuel Karlin, Protein length in eukaryotic and prokaryotic proteomes, Nucleic Acids Research, Volume 33, Issue 10, 1 June 2005, Pages 3390–3400, https://doi.org/10.1093/nar/gki615