Занятие 9. Исследование белок-нуклеиновых контактов
-
#1 Исследование контактов между молекулами белка и нуклеиновой кислоты
Для выполнения задания был скачан необходимая запись банка PDB и написан скрипт.
| Полярные | Гидрофобные | Всего | |
| Контакты белка с ... | |||
| ... остатками 2'-дезоксирибозы | 6 | 58 | 64 |
| ... остатками фосфорной кислоты | 21 | - | 21 |
| ... остатками азотистых оснований со стороны большой бороздки | 1 | 11 | 13 |
| ... остатками азотистых оснований со стороны малой бороздки | 2 | 3 | 5 |
Можно легко заметить, что наибольшее количество контактов образуется за счет гидрофобных взаимодействий с остатками 2'-дезоксирибозы. Контактов белка с остатками азотистых оснований в несколько раз меньше. Основную роль в этих взаимодействиях играют гидрофобные взаимодейтсвия оснований большой борозды. Малая борозда, видимо в силу пространственных затруднений контактирует с ДНК хуже.
#2 Последовательность
Доменная структура

Наиболее интереным остатком мне показался Лизин 706. Если посмотреть структуру, то видно, что именно после контакта с ним цитозина (в PDB файле цитозин 114 цепи P) происходит обрыв цепи. То есть, вероятно именно данный остаток является частью специфического сайта связывания, который ищет нужный праймер.
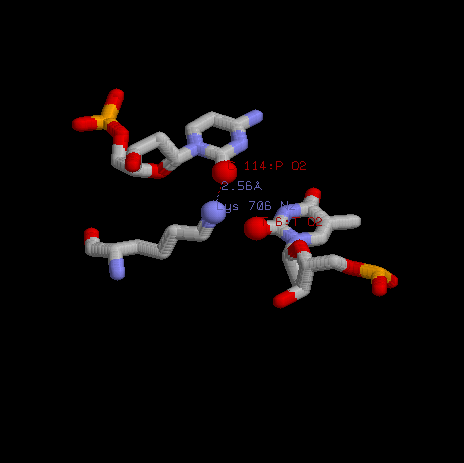 |
На данной иллюстрации можно проследить, что лизин пожет взаимодейтсвовать не только с цитозином цепи P, но и с комплементарным ему гуанином. |
 |
Здесь видно, что после контакта с лизином цепь P (оранжевая) прерывается. |
Предположительно, лизин 706 начинает расплетание цепочки ДНК. Это соответствет функции белка — это ДНК-полимераза. Белки этого семейства синтезируют цепь ДНК при наличии праймера и матричной цепи. Но как можно увидеть на выравнивании доменов Pfarm данная позиция не консервативна. Видимо этот лизин не играет значительной роли в работе полимеразы.
Name: DPOL_BPR69/391-859 Len: 26 Check: 5484 Weight: 1.00
Name: DPOL_BPT4/385-855 Len: 26 Check: 5393 Weight: 1.00
Name: DPOL_ASFL6/457-978 Len: 26 Check: 5168 Weight: 1.00
Name: DPOL_CBEPV/534-861 Len: 26 Check: 4863 Weight: 1.00
Name: DPOL_DESST/348-763 Len: 26 Check: 4882 Weight: 1.00
Name: DPOL_EBV/532-1013 Len: 26 Check: 5547 Weight: 1.00
//
1 26
DPOL_BPR69/391-859 AGPPLGSKGI GGFWTGKKRY ALNV..
DPOL_BPT4/385-855 SCPPLGSKGV GGFWKAKKRY ALNV..
DPOL_ASFL6/457-978 RGIDIIKQGQ TKLTKTIGTR IMEE..
DPOL_CBEPV/534-861 KGTALIRRDC TEIHKTILKN TIDI..
DPOL_DESST/348-763 RGLEIVRRDW SEIAKETQAR VLEA..
DPOL_EBV/532-1013 KGVELVRKTA CKFVQTRCRR VLDL..
Если посмотреть выдачу программы nucplot то можно заметить что она находит меньше связей, чем ручной анализ файла PDB. Во-первых, программа пренебрегает гидрофобными связями связями с сахарофосфатным остовом. Вероятно это это сделано умышленно, поскольку такие связи не отвечают за специфическое связывание, и, следоваетльно, не так интересно исследователям. Зато программа находит водородные связи посредством молекулой воды, которые выпадают из рассмотрения при ручном анализе. Интересно что часть цепи Т, не имеющей комплементарной, крайне насыщенна связями с белком, по сравнению с остальной цепью. Предположу что эта часть является частью расплетенной за счет этих контактов цепи.
{kind=link}