1. База данных OPM
Мне достался белок с pdb ID 3oz2. Этим белком оказался geranylgeranyl bacteriochlorophyll reductase-like из организма Thermoplasma acidophilum. Я попыталась найти его в базе данных OРM, и у меня не вышло. Я сделала вывод, что в этой базе белка нет, поэтому взяла себе другой с ID 3kcu. Это транспортер формиата (Formate transporter 1, FocA). Я взяла цепь А. Мембрану пронизывали в основном альфа-спирали, поэтому я нашла другой белок, где в мембране сидят бета-тяжи. Это белок 4mee - (Diffuse adherence adhesin). Данные по обоим белкам - в таблице.
| 3kcu | 4mee | |
| Толщинa гидрофобной части мембраны | 29.9 ± 0.7 Å | 26.4 ± 2.4 Å |
| Tрансмембранные спирали (тяжи) | 1(31-56), 2(65-85), 3(113-135), 4(161-181), 5(188-205), 6(210-219), 7(250-276) | 1(1006-1012), 2(1030-1040), 3(1048-1058), 4(1080-1087), 5(1098-1108), 6(1127-1138), 7(1153-1164), 8(1187-1198), 9(1211-1221), 10(1243-1253), 11(1260-1267), 12(1276-1284) |
| Среднее количество остатков в одной спирали | 19.86 | 9.25 |
| Мембрана | Внутренняя мембрана грамотриц. бактерии | Наружняя мембрана грамотриц. бактерии |
| Изображение в Jmol | 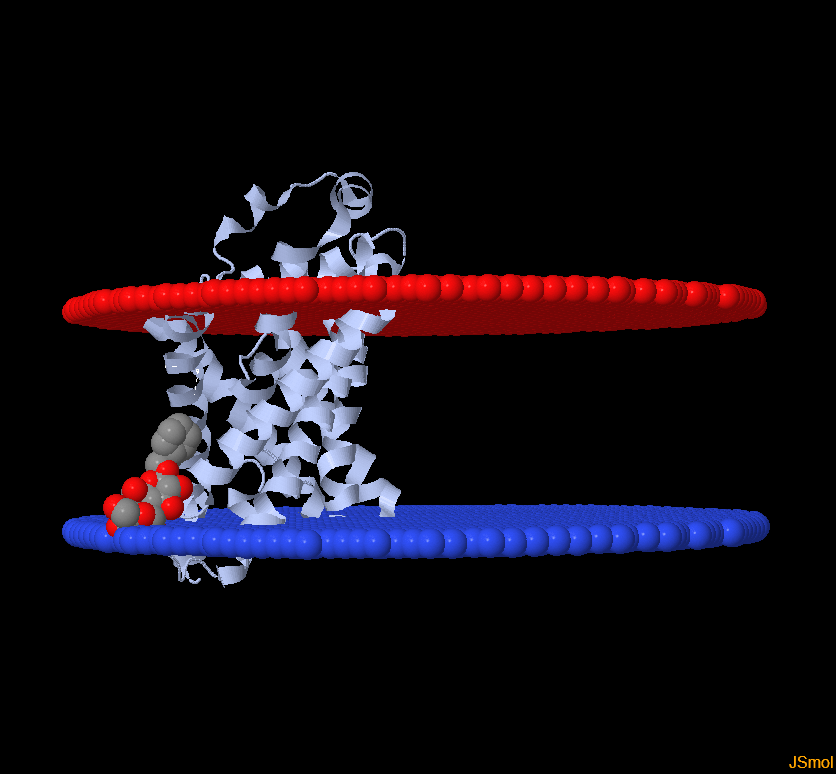 | 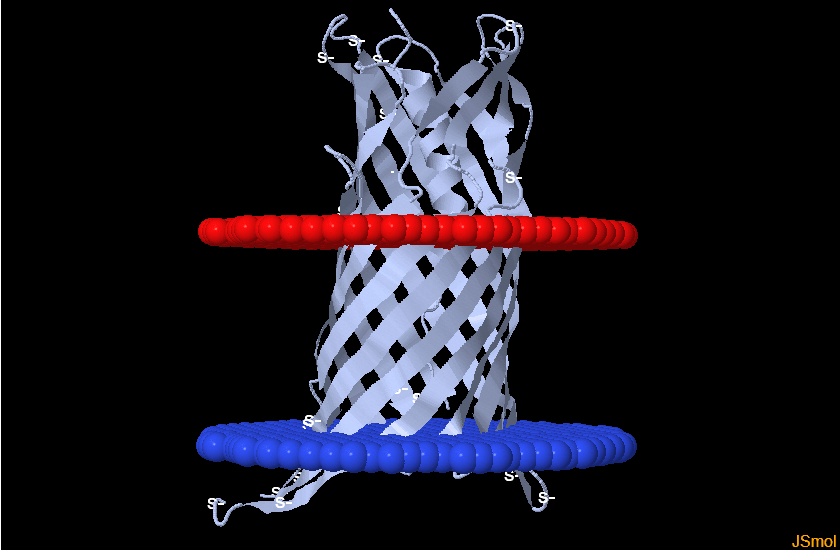 |
2. Анализ предсказания трансмембранных спиралей
Я запустила сервисы TMHMM и Phobius для белка 3kcu_A. Результат - в таблице:
| TMHMM | Phobius |
# pdb|3KCU|A Length: 285 # pdb|3KCU|A Number of predicted TMHs: 6 # pdb|3KCU|A Exp number of AAs in TMHs: 136.27998 # pdb|3KCU|A Exp number, first 60 AAs: 22.40287 # pdb|3KCU|A Total prob of N-in: 0.96583 # pdb|3KCU|A POSSIBLE N-term signal sequence pdb|3KCU|A TMHMM2.0 inside 1 34 pdb|3KCU|A TMHMM2.0 TMhelix 35 57 pdb|3KCU|A TMHMM2.0 outside 58 76 pdb|3KCU|A TMHMM2.0 TMhelix 77 99 pdb|3KCU|A TMHMM2.0 inside 100 118 pdb|3KCU|A TMHMM2.0 TMhelix 119 141 pdb|3KCU|A TMHMM2.0 outside 142 160 pdb|3KCU|A TMHMM2.0 TMhelix 161 183 pdb|3KCU|A TMHMM2.0 inside 184 194 pdb|3KCU|A TMHMM2.0 TMhelix 195 217 pdb|3KCU|A TMHMM2.0 outside 218 255 pdb|3KCU|A TMHMM2.0 TMhelix 256 278 pdb|3KCU|A TMHMM2.0 inside 279 285 |
ID pdb|3KCU|A FT TOPO_DOM 1 34 CYTOPLASMIC. FT TRANSMEM 35 55 FT TOPO_DOM 56 74 NON CYTOPLASMIC. FT TRANSMEM 75 98 FT TOPO_DOM 99 117 CYTOPLASMIC. FT TRANSMEM 118 141 FT TOPO_DOM 142 160 NON CYTOPLASMIC. FT TRANSMEM 161 180 FT TOPO_DOM 181 191 CYTOPLASMIC. FT TRANSMEM 192 212 FT TOPO_DOM 213 254 NON CYTOPLASMIC. FT TRANSMEM 255 277 FT TOPO_DOM 278 285 CYTOPLASMIC. // |
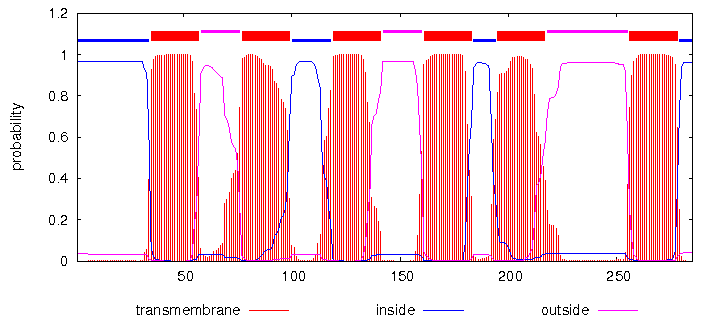 | 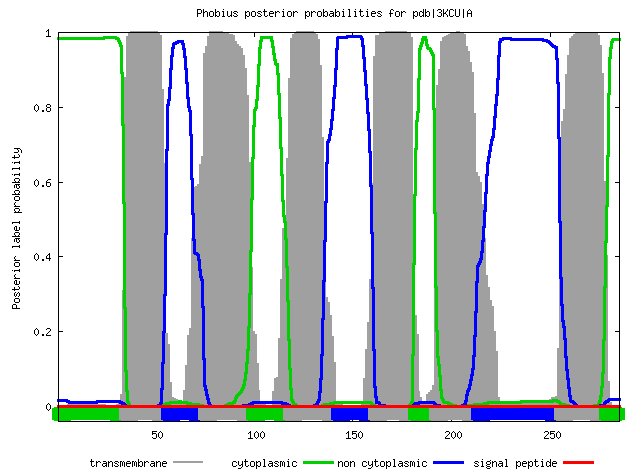 |
3. База данных TCBD
Я нашла в БД TCBD ,белки 3kcu и 4mee. TC-код белка 3kcu - 1.A.16.1.1. 1 - это значит канал, А - из альфа-спиралей, 16 - семейство FNT (транспортеры формиата-нитрита), 1.1 - непосредственно указывает на белок. Вот что написано в TCBD про это семейство:
Члены семьи FNT были секвенированы из грам-отрицательных и грам-положительных бактерий, архей и дрожжей. Прокариотические белки семейства FNT предположительно учавствуют в транспорте структурно родственных соединений, формиатов и нитритов. За исключением дрожжевого белка (627 аминокислотных остатков), все члены семьи имеют длину 256-285 остатков и образуют 6-8 предполагаемых трансмембранных α-спиральных домена (ТМS). В одном случае, в белке E. coli FocA, была создана топология из 6 TMS. Дрожжевой белок имеет похожую топологию, но помимо этого большой с-концевой гидрофильный хвост около 400 остатков. Филогенетическое дерево показывает кластеризацию в зависимости от функции и организменной филогении. Бактериальные FocA, связанные с pfl, образуют кластер I; FdhC бактерий и архей, связанные с формиат-дегидрогеназы составляют кластер II; NirC бактерий относятся к кластер III, и дрожжевой белок образует кластер IV. Энергетические механизмы связывания для белки семейства FNT не были подробно охарактеризованы. Захват HCO2 - и NO2, вероятно, связан с симпортом протонов. HCO2 - высвобождение может быть вызвано мембранным потенциалом с помощью унипорт механизма или H антипорта. FocA Е.coli катализирует двунаправленный транспорт формиата, имеет пентамерную четвертичную структуру и может работать по канальному механизму. FocA, типичный представитель FNT, транспортирует короткоцепочечные кислоты у бактерий, архей, грибов, водорослей и некоторых эукариотических паразитов. Wang и соавт. (2009) сообщили, что кристаллическая структура FocA кишечной палочки в разрешении 2.25 образует симметричный пентамер, с каждым промотером состоящим из шести ТMS. Несмотря на отсутствие гомологии последовательности, общая структура промотера FocA напоминает промотер аквапорина, указывая, что FocA - это канал, а не перевозчик. Структурный анализ выявлил потенциально важные остатки канала, определил траекторию канала и выявил два сайта констрикции. В отличие от аквапорина, FocA непроницаем для воды, но обеспечивает прохождение формиата. FocA (2.А.44.1.1) может переключить режим работы с пассивного экспортного канала при высокой рН на вторичный активный формиат/H импортер при низких значениях рН. Кристаллическая структура FocA Salmonella typhimurium при рН 4.0 показывает, что этот параметр включает в себя серьезную перестановку амино-концов отдельных промотеров в пентамерном канале. Амино-концевые спирали открывают или блокируют транспорт в согласованно и совместно, что показывает, как FocA рН-зависим. Электрофизиологические исследования показывают, что белок выступает в качестве специфического формиат канала при рН 7.0 и что он закрывается на сдвиг рН до 5,1. Вероятные транспортные реакции, катализируемые различными членами семьи FNT являются: (1) RCO2- or NO2- (out) = RCO2- or NO2- (in) (2) HCO2- (in) = HCO2- (out) (3) HS- (out) = HS- (in)Что касается второго белка, 4mee, его TC-код 1.B.12.1.1. Этот белок относится к каналам (1) из бета-тяжей (В), семейству автотранспортеров (12).