2.Кристалл из белков
Для структуры белка Pol μ (PDB ID= 5TYB) в файле PDB записанны параметры элементраной ячейки кристалла:
CRYST1 60.047 68.488 110.450 90.00 90.00 90.00 P 21 21 21 4
Первые три числа (60.047; 68.488; 110.450) означают длинны направляющих векторов в Å. За ними следуют углы между векторами (90.00; 90.00; 90.00), которые для данной структуры равны 90°. P 21 21 21 - тип симмметрии кристалла по символике Германа-Моргена. В случае структуры 5TYB это примитивная группа (P), с двумя осями винтовой симметрии типа 21 и осью повортной симметрии типа 21. Последняя цифра отражает количество молекул в ячеке - 4.
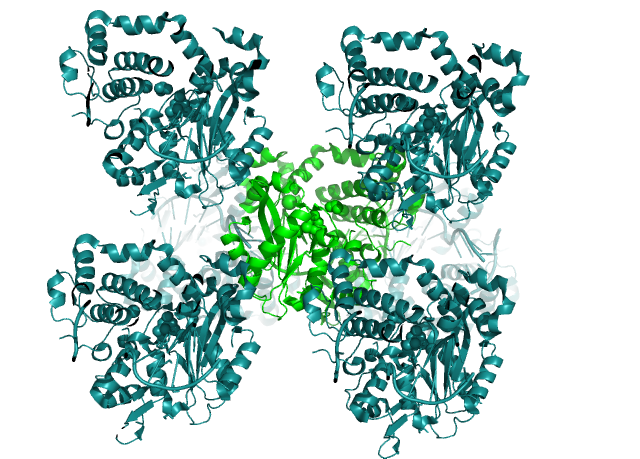
Рисунок 1. Востоновленная кристалическая структура белка Pol μ (PDB ID= 5TYB ) в пределах 5Å.
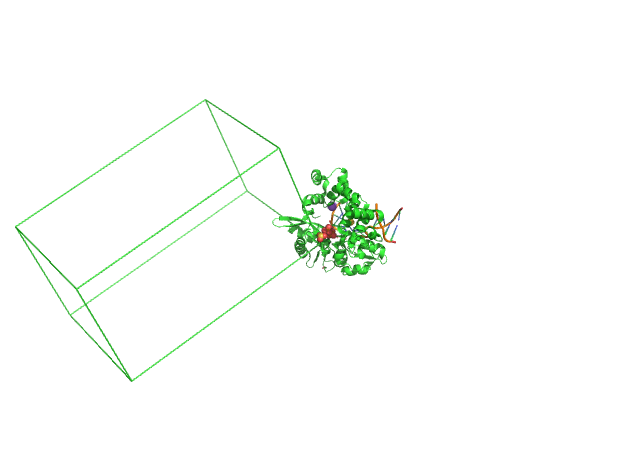
Рисунок 2. Расположение исходной молекулы относительно элементарной ячейки.
На рис. 2 видно, что исходная молекула не включена в элементарную ячеку, что является нежелательно для PDB структур, хотя явной ошибки тут нет.
Анализ контактов
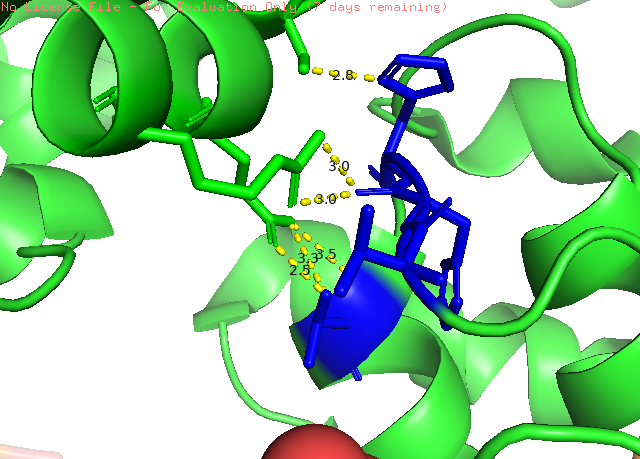
Рисунок 3. Пример полярных взаимодейтсвий между мономерами в кристалле белка Pol μ. Показанно взаимодействие пептидных цепей. Зеленным показанна ассиметрическая еденица, а синим - окружение. Желтым пунктиром обозначенна водородная связь (точнее линия от донора до акцептора), с указанием длинны в Å.
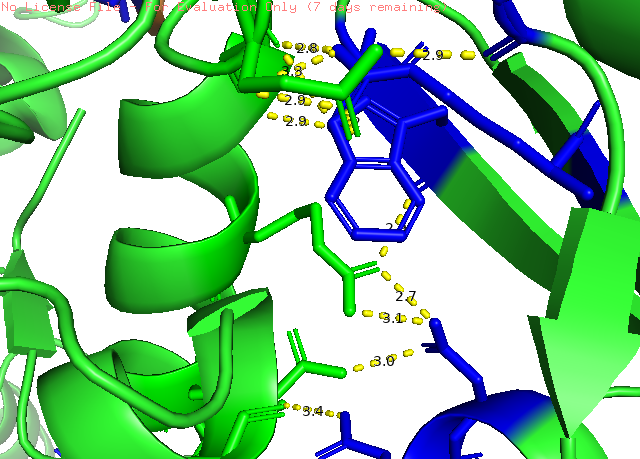
Рисунок 4. Пример полярных взаимодейтсвий между мономерами в кристалле белка Pol μ. Показанно взаимодействие пептидных цепей. Обозначение аналогично рис. 3.
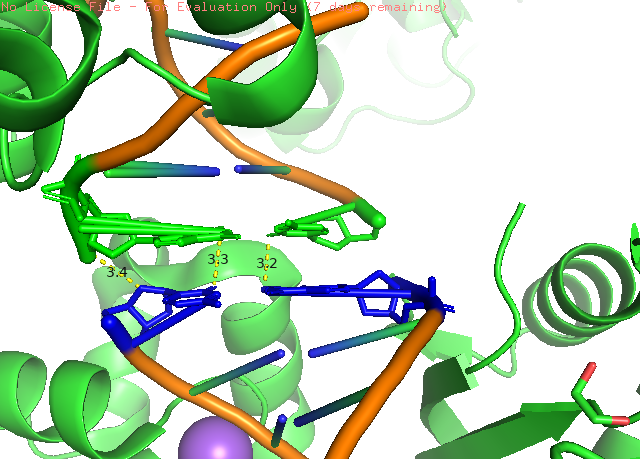
Рисунок 5. Пример полярных взаимодейтсвий между мономерами в кристалле белка Pol μ. В данном примере показанно взаимодейтсвие нуклеиновых кислот. Обозначение аналогично рис. 3.
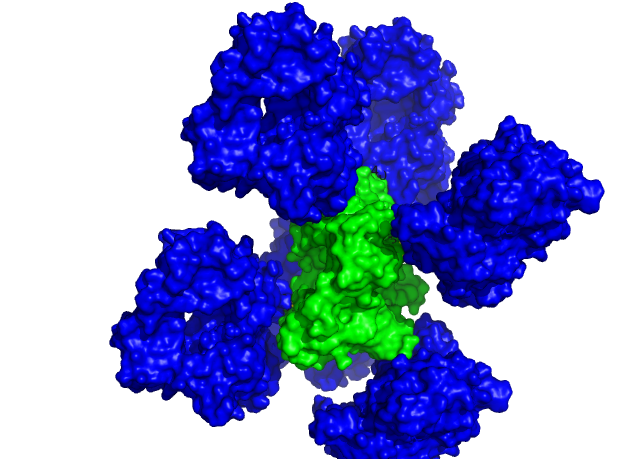
Рисунок 6. Общий вид на взаимодейстие ассиметрической едениы с окружением. Обозначения прежние.
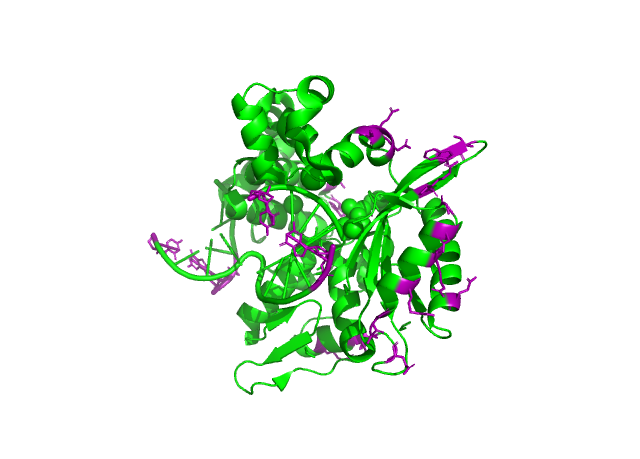
Рисунок 7. Зоны контактов ассиметрической еденицы с окружением, cartoon. Остатки, приниаюие участие в контакте показанны фиолетовым.
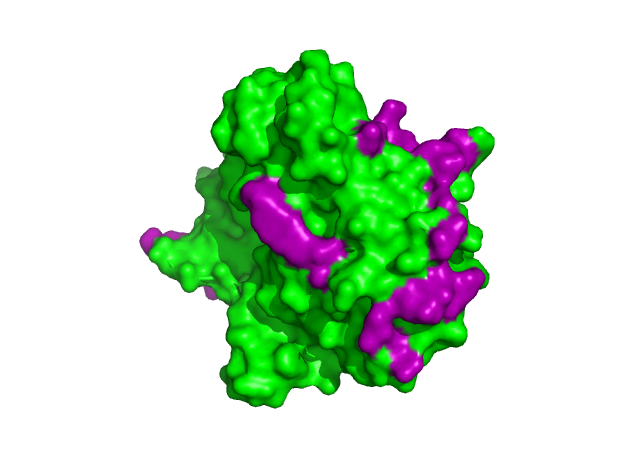
Рисунок 8. Зоны контактов ассиметрической еденицы с окружением, поверхность. Остатки, приниаюие участие в контакте показанны фиолетовым.
Всего я обнаружил 6 зон контактов, гидрофобных взаимодействий обнаруженно не было. Однако я считаю маловероятно что хотя бы одна из зон принимает участие в образовании олигомера, в следствии специфичности субстрата, а именно поврежденной двухцепочечной молекулы ДНК. Скорее всего, при удлиненнии субстрата, будет нарушаться кристалообразование, т.е. при условиях приближенным к in vivo 2 зоны котакта связанные с ДНК точно исчезнут, а оставшиеся зоны не несут явного эволюционного приемущества. Но все же полностью убирать из расммотрения возможность образования олигомеров я бы не стал.
Странное расположение белковых цепей в структуре 1mnm
Если открыть структуру 1mnm (комплекс блок-ДНК), то может насторожить, каким образом где-то в стороне от основной молекулы образовался одинокий бета-стренд? Действительно, глядя на рис. 9 это никак не очевидно.
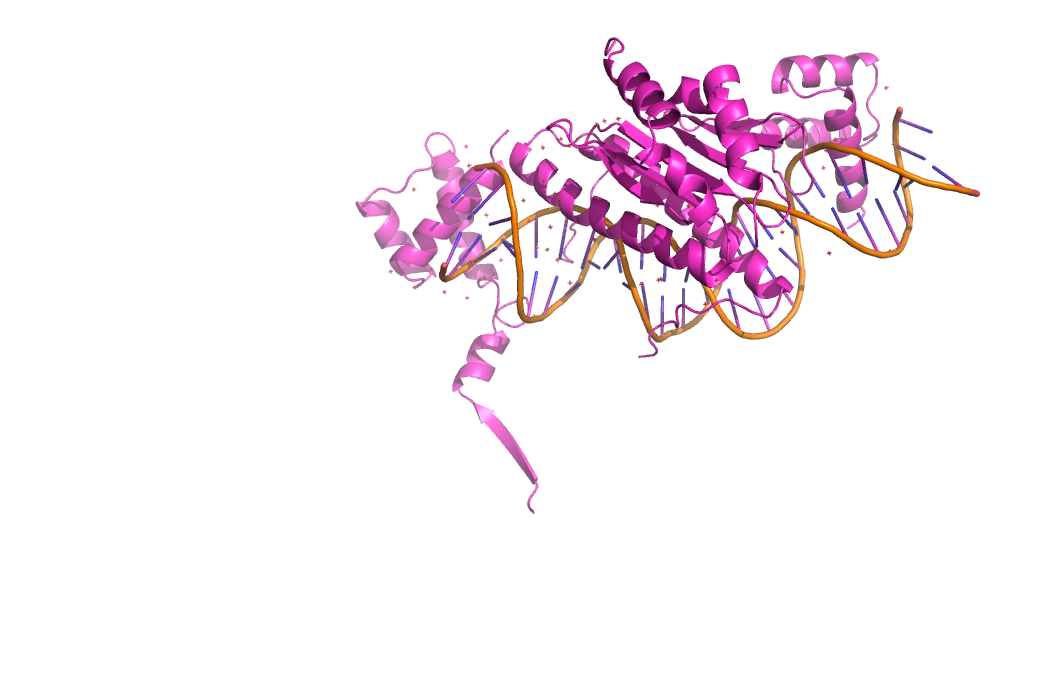
Рисунок 9. Сруктура 1mnm.
Однако, стоит лишь востоновить кристалическую решетку, как картина проясняется:
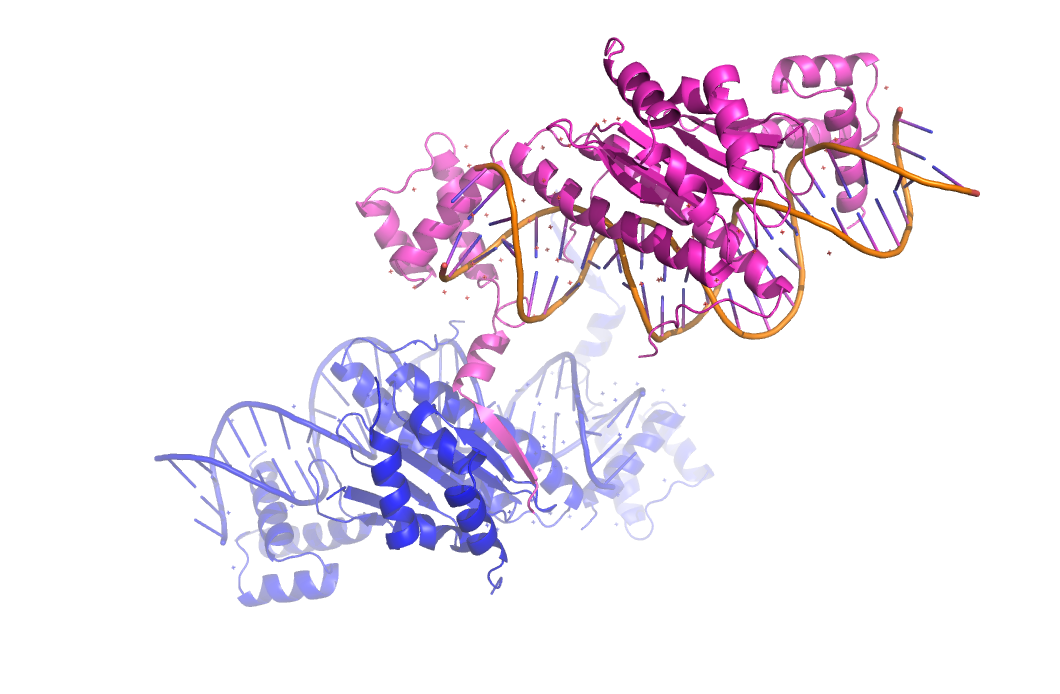
Рисунок 10. Востоновленная структура кристала 1mnm. Показанно образование бета-стренда между исходной (фиолетовый) и соседней (синий) молекулами белка в кристалле.
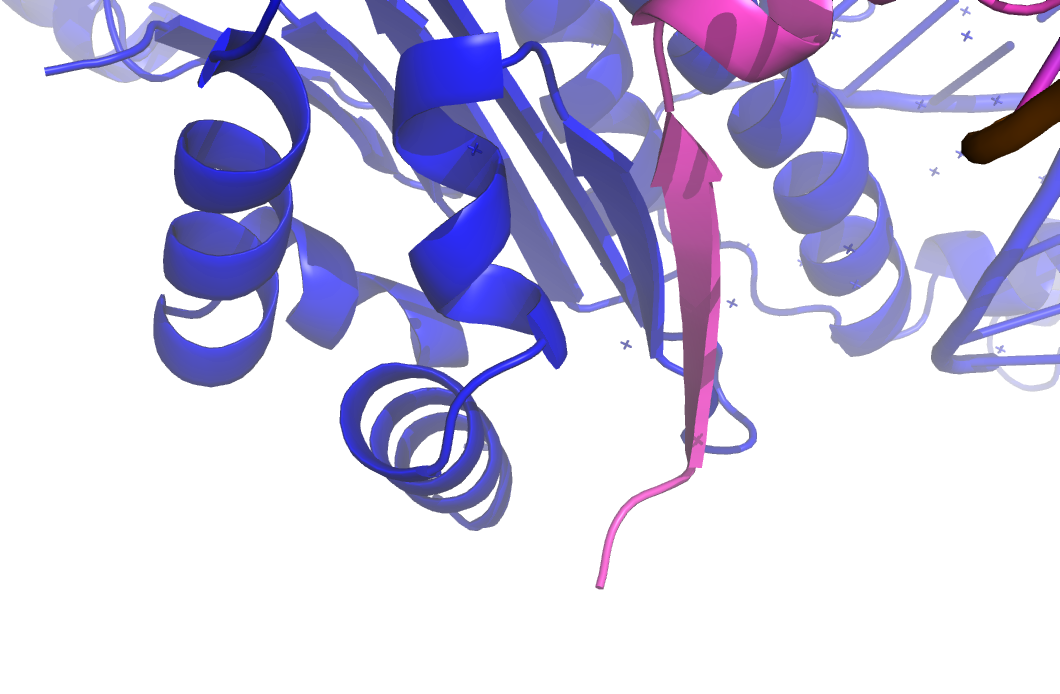
Рисунок 11. Более близкий взгляд на контакт с цепочкой белка в соседей ячейке.
Т.е. такое странное явление очень просто обьясняется контактами между соседними белковыми цепями в кристалле, о которых, конечно же, никогда не стоит забывать.
Примеры PDB структур, с несовпадением асимметрической и биологической единиц
У структуры 6bdc (белок Hcp1 бактерии Flavobacterium johnsoniae) 1 ассиметрическая еденица, в то время как бологическая состоит из 7, что отлично видно на рис. 12 и 13
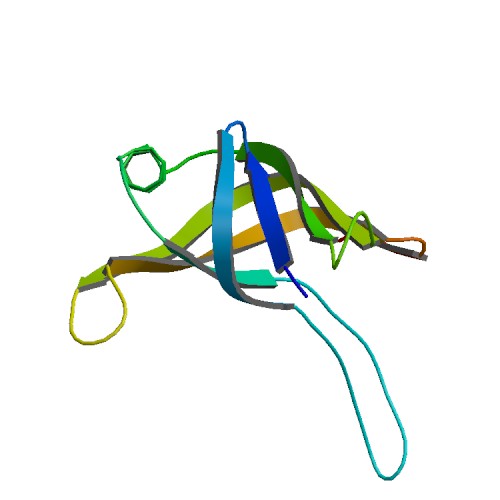
Рисунок 12. Структура 6bdc, ассиметрическая еденица.
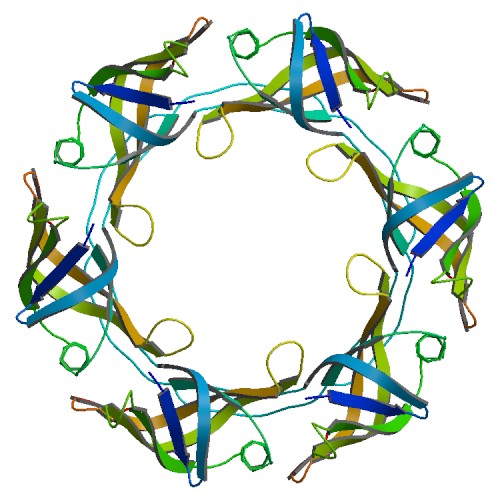
Рисунок 13. Структура 6bdc, биологическая еденица, состоящая из 7 ассиметрических.
Для 5YSO (Estrogen Related Receptor-3 или иначе ERR-gamma) (рис. 14) ситуация обратная - 1 ассиметрическая еденица содержит аж 3 бологические.
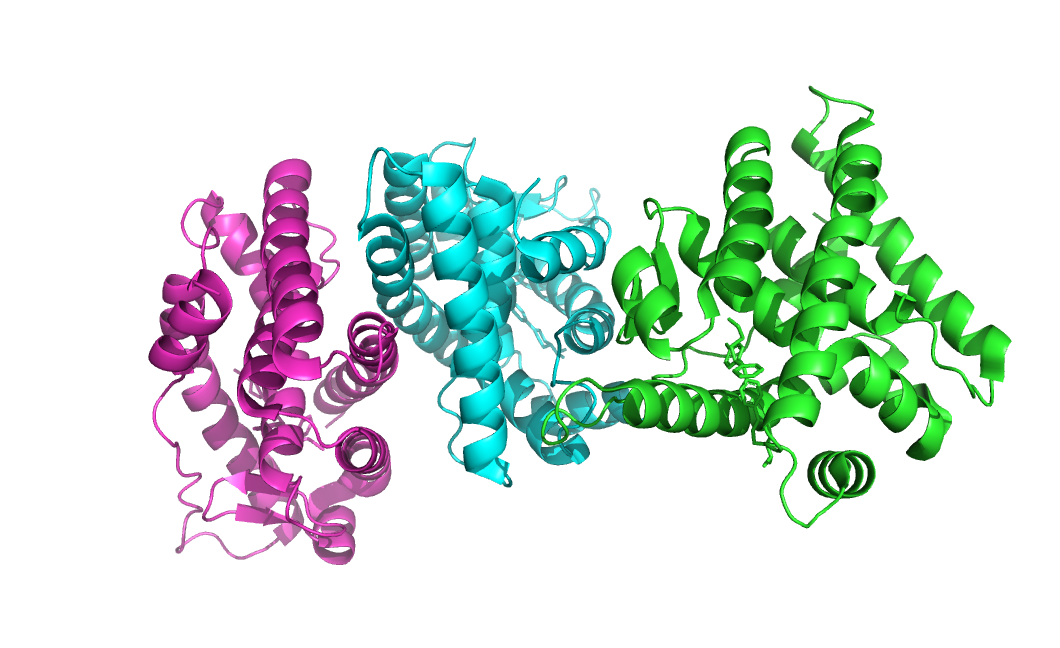
Рисунок 14. Ассиметрическая еденица структуры 5YSO, бологигические еденицы покрашенны в разные цвета.
| на главную |
© Гавриш Глеб 2017 |