Задание 1. Предсказание вторичной структуры заданной тРНК
В данном практикуме с помощью команды einverted из пакета EMBOSS мне удалось найти инвертированные участки в нуклеотидных последовательностях и возможные комплементарные участки в последовательности исследуемой тРНК 1EIY.
Была использована команда: einverted -sequence 1eiy.seq -gap 12 -threshold 10 -match 3 -mismatch -3 -outfile outfile -outseq seqout
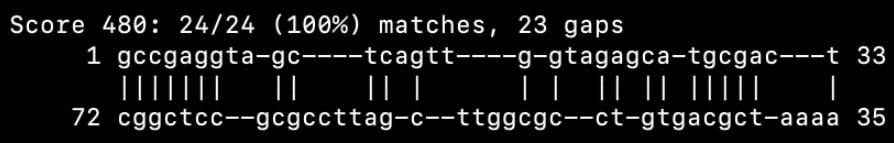
Рис. 1 Выдача программы einverted из пакета EMBOSS.
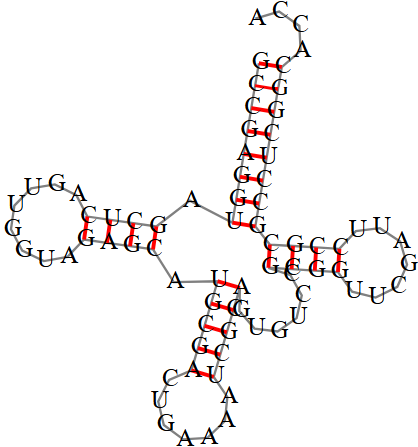
Рис. 2 Cтруктура, предсказанная алгоритмом Зукера, для 1EIY.
| Участок структур | Позиции в структуре | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
|---|---|---|---|
| Акцепторный стебель | 5'-1-7-3' 5'-66-72-3' | 5'-1-7-3' 5'-66-72-3' | 7 |
| D-стебель | 5'-10-13-3' 5'-22-25-3' | - | 4 |
| Т-стебель | 5'-49-53-3' 5'-61-65-3' | - | 5 |
| Антикодоновый стебель | 5'-27-31-3' 5'-39-43-3' | 5'-27-31-3' 5'-39-43-3' | 5 |
| Общее число неканонических пар нуклеотидов | 22 | 12 из 24 предсказано верно | 21 |
Таблица 1. Реальная и предсказанная вторичная структура тРНК из файла 1EIY.pdb
Задание 2. Поиск ДНК-белковых контактов в заданной структуре
Упражнение 1.
Для анализа был взят комплекс ДНК и белка 1hw2.pdb. Был написан скрипт на Pymol для выделения следующих множеств атомов: 1) set 1- множество атомов кислорода 2'-дезоксирибоз, 2) set 2- множество атомов кислорода в остатке фосфорной кислоты, 3) set 3- множество атомов азота в азотистых основаниях.
Далее был написан второй скрипт, благодаря котрому было получено последовательное изображение всей структуры, но с выделенными шариками множеством атомов set1, затем set2 и set3
Упражнение 2.
| Контакты атомов белка с | Полярные | Неполярные | Всего |
|---|---|---|---|
| Остатками 2'-дезоксирибозы | 7 | 38 | 45 |
| Остатками фосфорной кислоты | 12 | 26 | 38 |
| Остатками азотистых оснований со стороны большой бороздки | 11 | 13 | 24 |
| Остатками азотистых оснований со стороны малой бороздки | 3 | 4 | 7 |
Таблица 2. Контакты разного типа в комплексе 1HW2.pdb
На основе данных из таблицы можно сделать вывод:
1. Остатки 2'-дезоксирибозы и фосфорной кислоты в основном образуют неполярные контакты
2. Остатки азотистых оснований со стороны большой и малой бороздок образуют почти одинаковое количество полярных и неполярных контактов
3. Со стороны большой бороздки гораздо больше взаймодействий, чем со стороны малой
Упражнение 3.
Используя программу nucplot, была получена схема ДНК-белковых контактов.
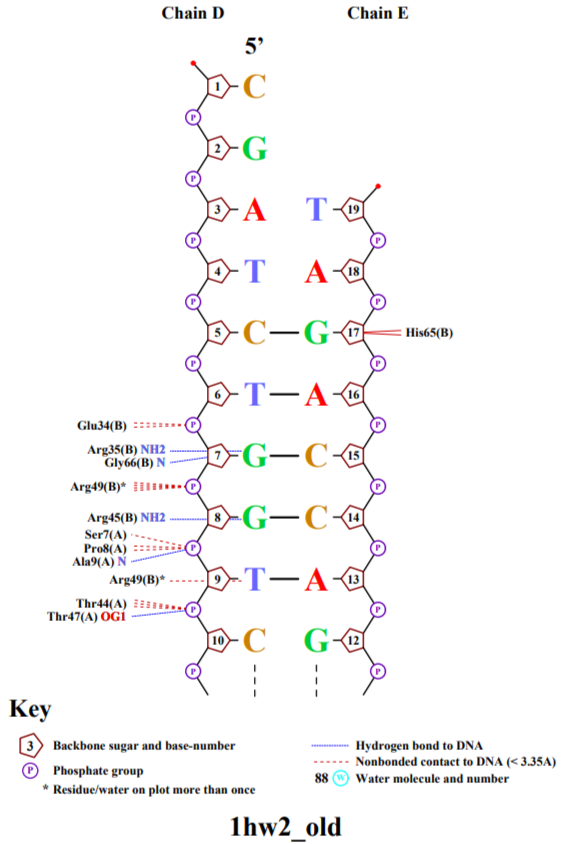
Рис. 3-4 Cхема ДНК-белковых контактов.
Упражнение 4.
Среди взаимодействующих аминокислот наибольшее число контактов с ДНК имеет 65 гистидин цепи A белка. Он взаимодействует с 16 аденином цепи D и остатком дезоксирибозы 17 гуанина цепи D.
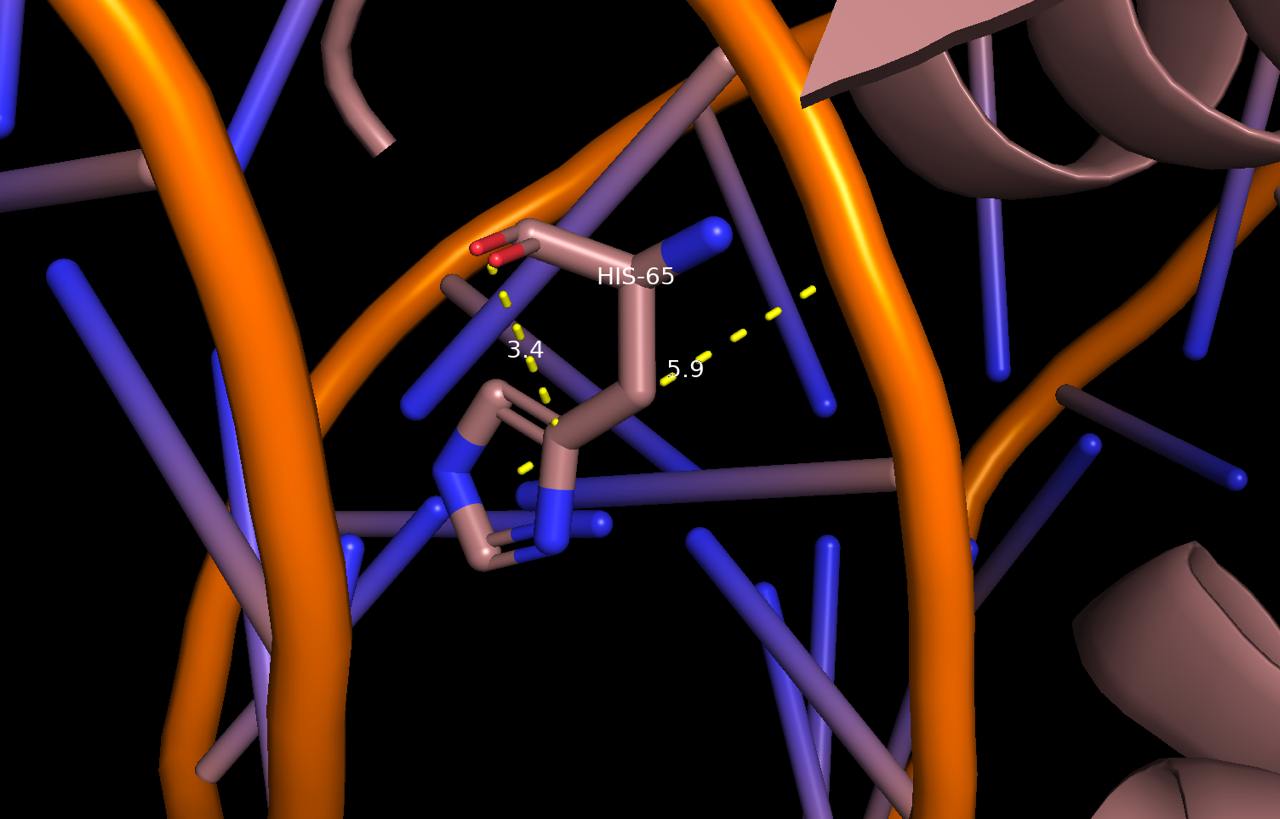
Рис. 5 Взаимодействие His65
Наиболее важным для рапознавания ДНК является 49 аргенин на цепи В белка, взаимодействующий с 9 тимином цепи D и атомом фосфора. Аминокислотный остаток проникает в ДНК.
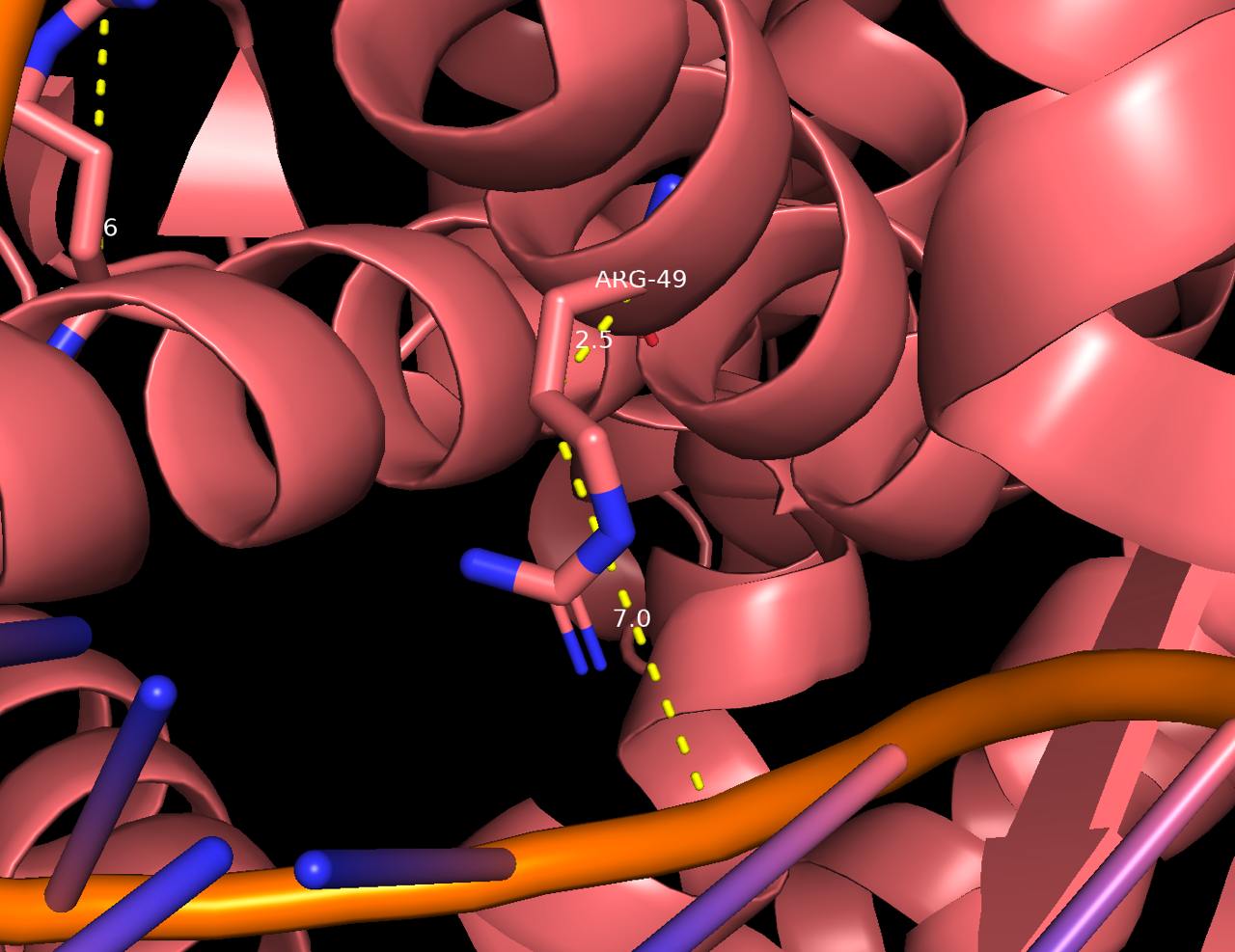
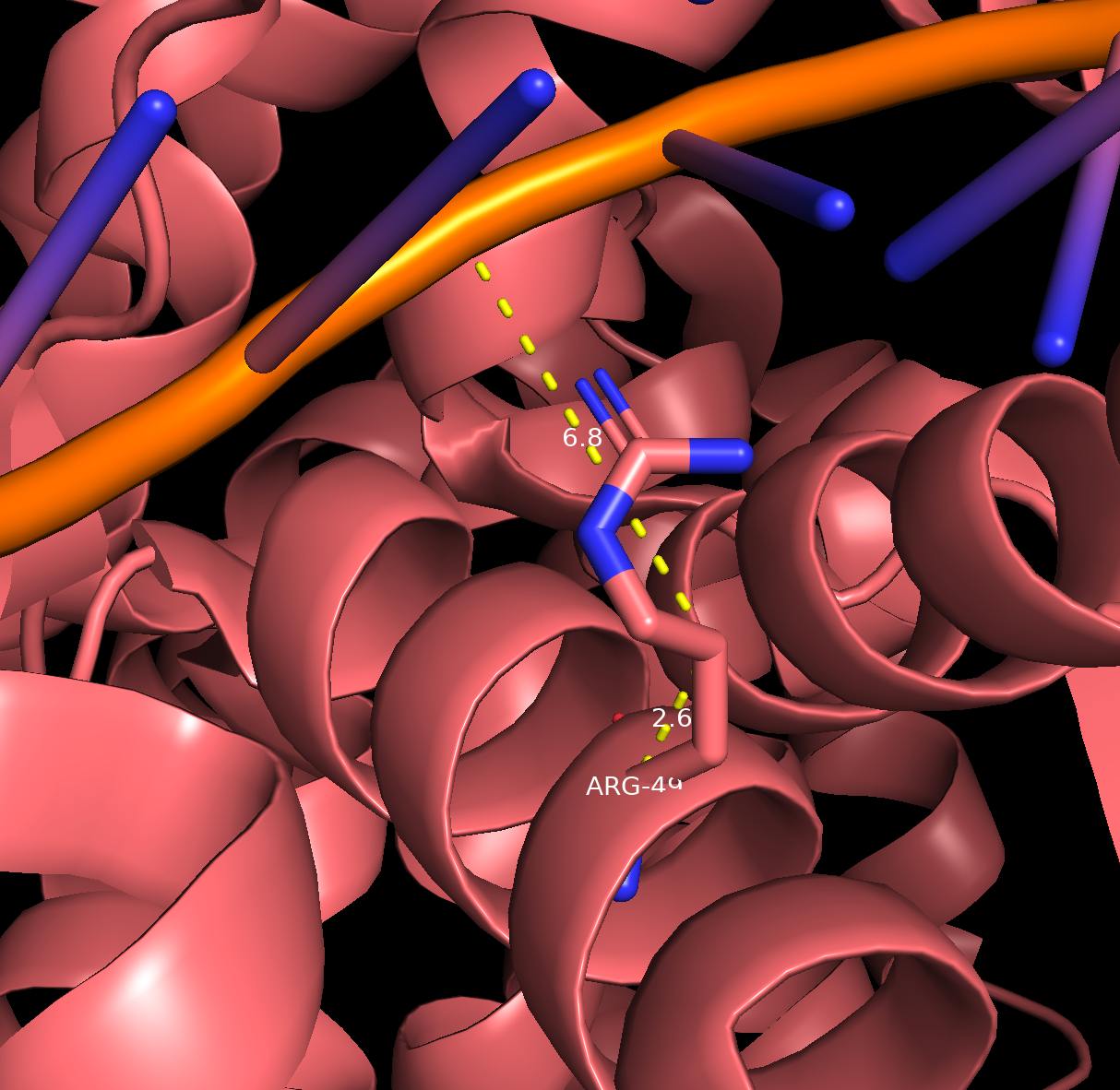
Рис. 6 Взаимодействие Arg49