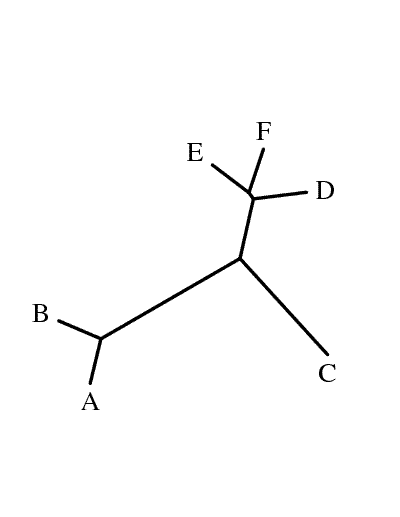
Для визуализации заданного скобочной формулой дерева можно использовать программу drawgram из Online-версии пакета Phylip.
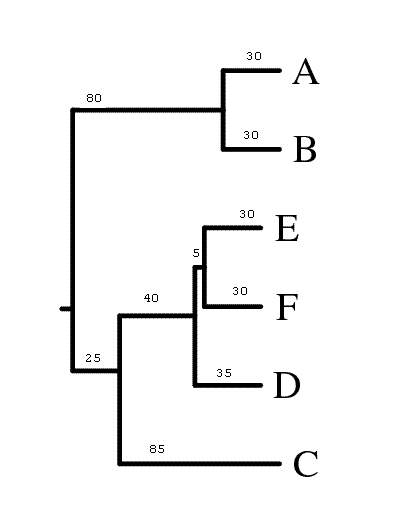
Описание разбиений оформлено в виде таблички, столбцы которой соответствуют листьям дерева, а строки - ветвям дерева. Описание ветвей, отделяющих один лист от всех остальных, опущено ввиду своей тривиальности.
| A | B | C | D | E | F |
| 1 | 1 | 0 | 0 | 0 | 0 |
| 1 | 1 | 1 | 0 | 0 | 0 |
| 1 | 1 | 1 | 1 | 0 | 0 |
Ген имеет длину 780 н.о. (259 а.о. белка *3 + 1 стоп-кодон). Мутацию в стоп-кодоне будем считать равнозначной мутации в любом другом кодоне (на самом деле стоп-кодон не сохранился ни в одном из листьев дерева, но если бы и сохранился, работоспособность полученных в ходе нашей "случайной" эволюции белков маловероятна).
Формула для пересчёта расстояний M в число мутаций в гене N: N=M*780/100
Для получения "мутантов" используется программа msbar пакета EMBOSS. Она делает в гене заданное число замен, причем нуклеотид заменяется на любой из четырех (а не трех остальных). Поэтому четверть замен заменами не являются и программе на вход необходимо задавать в 4/3 раз больше замен, т.е. N =4*M*780/(3*100) =10.4*M
Формула для пересчёта расстояний M в значение параметра -count программы msbar N': N'=10.4*M
Скрипт для получения мутантных последовательностей:
msbar DeoC_gene.fasta AB.fasta -point 4 -count 832 -auto msbar DeoC_gene.fasta CDEF.fasta -point 4 -count 260 -auto msbar CDEF.fasta DEF.fasta -point 4 -count 416 -auto msbar DEF.fasta EF.fasta -point 4 -count 52 -auto msbar AB.fasta A.fasta -point 4 -count 312 -auto msbar AB.fasta B.fasta -point 4 -count 312 -auto msbar CDEF.fasta C.fasta -point 4 -count 884 -auto msbar DEF.fasta D.fasta -point 4 -count 364 -auto msbar EF.fasta E.fasta -point 4 -count 312 -auto msbar EF.fasta F.fasta -point 4 -count 312 -auto
Помещаем последовательности, соответствующие листьям дерева, в один файл в fasta-формате (он является "самым правильным" выравниванием, поскольку в ходе "эволюции" производились только замены и положения н.о. в последовательностях не изменялись).
- Реконструировать дерево алгоритмом максимального
правдоподобия (программа fdnaml):
fdnaml ali.fasta -ttratio 0.5 -auto
Параметр -ttratio равен 0.5, т.к. программа msbar делает все замены с одинаковой вероятностью и отношение числа транзиций к числу трансверсий равно 1/2. - Чтобы реконструировать дерево алгоритмами UPGMA или Neighbor-joining,
сначала надо посчитать попарные расстояния между последовательностями
программой fdnadist:
fdnadist ali.fasta -ttratio 0.5 -auto
Результат в виде файла с расширением .fdnadist подается на вход программе fneighbor. Команда:fneighbor abcdef.fdnadist -auto -outfile NJ.fneighbor
для реконструкции алгоритмом Neighbor-joining иfneighbor ali.fdnadist -treetype u -auto -outfile UPGMA.fneighbor
для реконструкции алгоритмом UPGMA.
Полученные деревья:
| Алгоритм | Дерево | Скобочная структура | Ветви (ABCDEF) | ||
| 110000 | 111000 | 111100 | |||
| Нет (исходное дерево) | См. выше | ((А:30,В:30):80,(((Е:30,F:30):5,D:35):40,С:85):25); | + | + | + |
| UPGMA |
+---------A
+-------------------------2
! +---------B
--5
! +--------------------------C
+--------4
! +----------D
+---------------3
! +--------E
+-1
+--------F |
((A:32.8,B:32.8):85.9,(C:87.9,(D:35.9,(E:29.7,F:29.7):6.2):52.0):30.8); | + | + | + |
| Neighbor-joining |
+--B ! ! +-------------C 1---------------------2 ! ! +-----D ! +------------3 ! ! +-------E ! +--4 ! +---F ! +---------A |
(B:15.6,(C:73.0,(D:28.9,(E:39.9,F:19.6):13.2):64.6):109.3,A:49.9); | + | + | + |
| Максимального правдоподобия |
+----B | | +-------D | +-----------4 | | | +------E 1--------------------2 +--3 | | +----F | | | +--------------C | +-------A |
(B:24.594,((D:38.7,(E:36.7,F:23.2):4.0):62.1,C:76.9):103.1,A:40.6); | + | + | + |
Сравнение деревьев
Все деревья имеют одинаковую топологию. Исходное дерево не было ультраметрическим, но посроенное по алгоритму UPGMA ультраметрическое дерево по длинам ветвей очень близко к исходному. В построенных по алгоритмам Neighbor-joining и максимального правдоподобия деревьях расстояния отличаются от исходных сильнее, в парах A-B и E-F листья A и E эволюционировали заметно быстрее листьев B и F (особенно в дереве NJ).Этапы работы:
- Программой fseqboot создано 100 бутстреп-реплик
выравнивания:
fseqboot ABCDEF.fasta -auto
- Полученные 100 выравниваний подаются на вход программе fdnaml. В
выходном файле (abcdef.treefile) окажется 100 скобочных формул,
соответствующих реконструкциям, сделанным по каждому из выравниваний.
fdnaml abcdef.fseqboot -ttratio 0.5 -auto
- Программа fconsense проводит бутстреп-анализ.
fconsense abcdef.treefile
Консенсусное дерево совпало по топологии с реальным деревом, но одна из внутренних ветвей встречается только в 41 дереве из 100:
Sets included in the consensus tree Set (species in order) How many times out of 100.00 cfedba ....** 100.00 .***.. 85.00 .**... 41.00Внутренняя ветвь ABCD-EF оказалась недостоверной, первые две из ветвей-аутсайдеров - конкуренты именно этой ветви:
Sets NOT included in consensus tree: Set (species in order) How many times out of 100.00 .*.*.. 29.00 ..**.. 18.00Такую недостоверность можно объяснить малой длиной этой ветви в исходном дереве (5 мутаций на 100 нуклеотидов). Две другие внутренние ветви и все внешние достаточно достоверны (встречаются более 70 раз). Неукорененное дерево:
+---------------------------C
|
| +------A
| +-------100.0-|
| | +------B
+------|
| +------F
| +-41.0-|
+-85.0-| +------E
|
+-------------D
Скобочная структура:
(C:100.0,((A:100.0,B:100.0):100.0,((F:100.0,E:100.0):41.0,D:100.0):85.0):100.0);(в скобках - достоверность ветви).
- Поместите скобочную формулу в отдельный файл.
- Подать файл на вход fdrawtree.
- Результат перевести в GIF-формат.