Вторичная структура РНК
С помощью
einverted -gap 12 -threshold 10 -match 3 -mismatch -3 -sequence 1i9v.fa -outfile outfile -outseq seqout
нашла инвертированные участки последовательности.
С помощью алгоритма Зукера (RNAfold) построила возможную структуру тРНК.
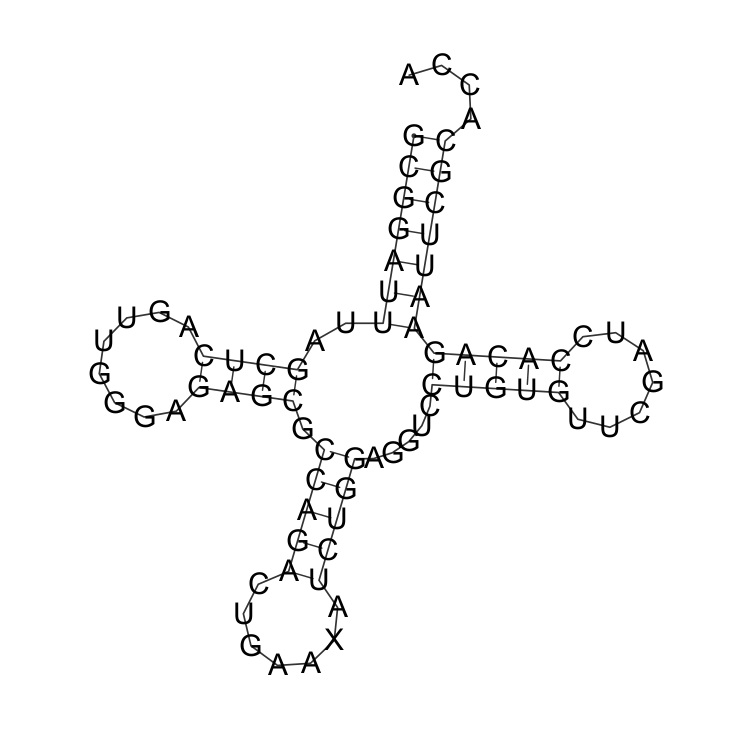
Сравнение вторичной структуры:
| Участок | Позиции в find_pairs | Результаты einverted | Результаты по алгоритму Зукера |
|---|---|---|---|
| Акцепторный стебель | 1-7:66-72//7 пар, 6 канонических | - | 7 из 7 пар |
| T-стебель | 49-52:62-65//5 пар | 5 из 5 пар | 5 из 5 пар |
| D-стебель | 10-13:22-25//4 пары | - | 4 из 4 пар |
| Антикодоновый стебель | 40-43:27-30//4 пары | 4 из 4 пар, есть 4 лишних | 4 из 4 пар, есть 1 лишняя |
| Общее число каноничсеких пар нт | 19 | 9 предсказано | 19 предсказано |
ДНК-белковые контакты
Скрипт в JMol, который покажет ДНК-белковые контакты
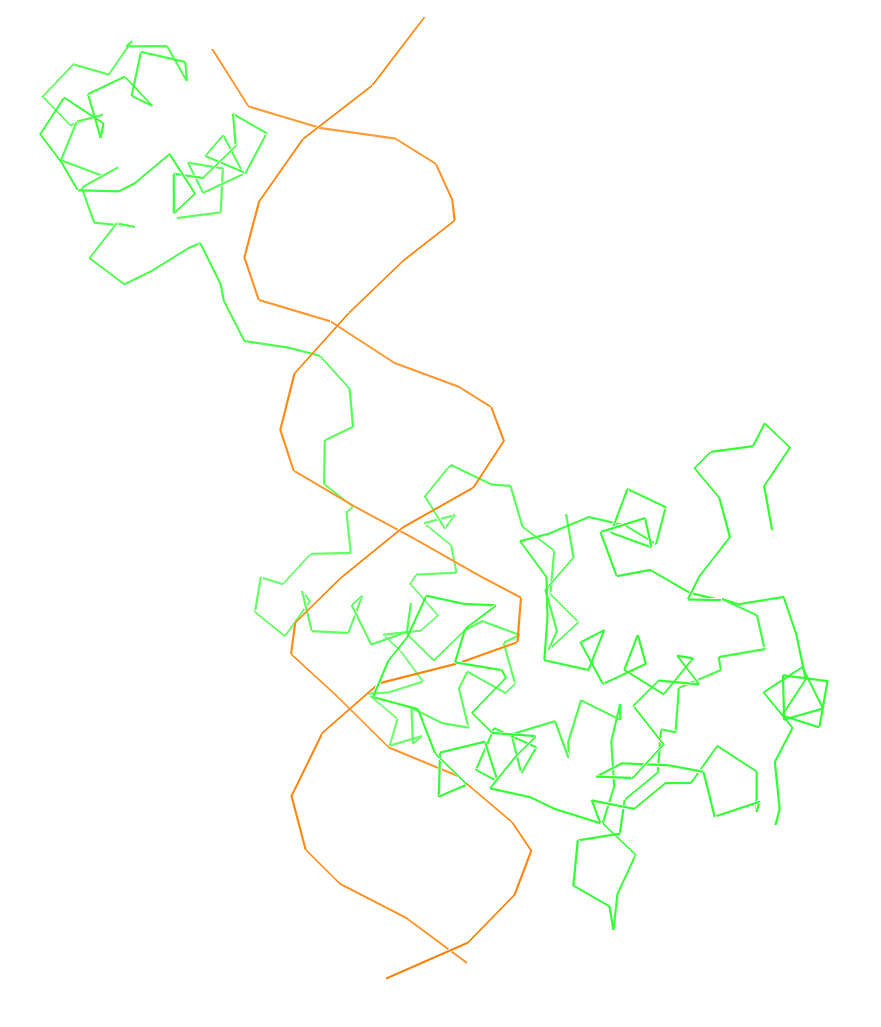
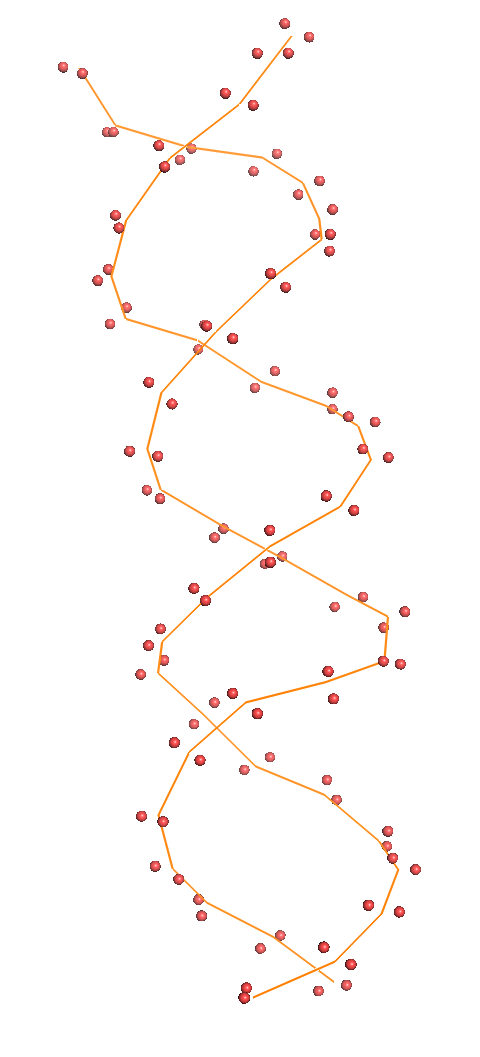
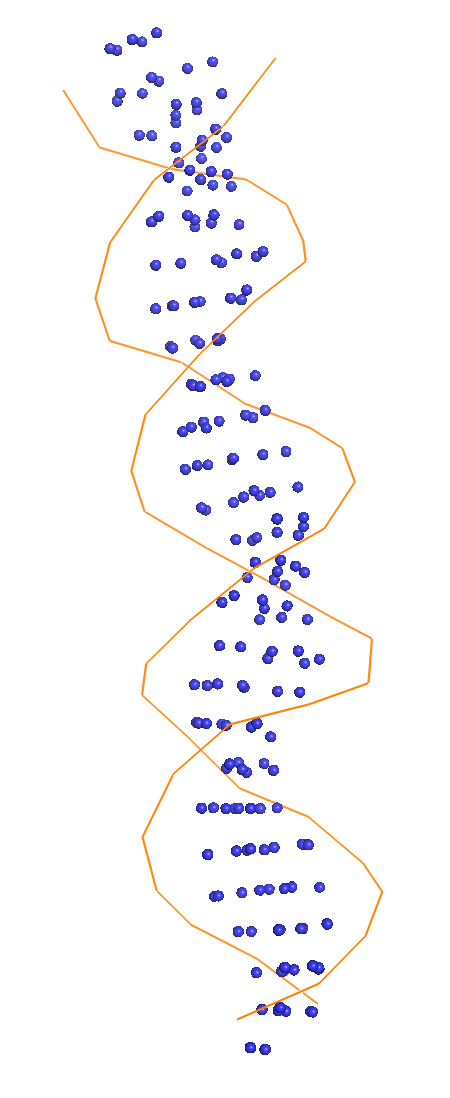
Весь комплекс//сахарофосфатный остов ДНК//set1//set2//set3
Результат запуска:
| Контакты атомов белка с | Полярные | Неполярные | Всего |
|---|---|---|---|
| остатки 2'-дезоксирибозы | 3 | 13 | 16 |
| остатки фосфорной кислоты | 9 | 6 | 15 |
| остатки азотистых оснований (большая бороздка) | 4 | 11 | 15 |
| остатки азотистых оснований (малая бороздка) | 0 | 1 | 1 |
Так же контакты можно описать с помощью nucplot. Исследуем цепь белка В и цепли ДНК C, D.
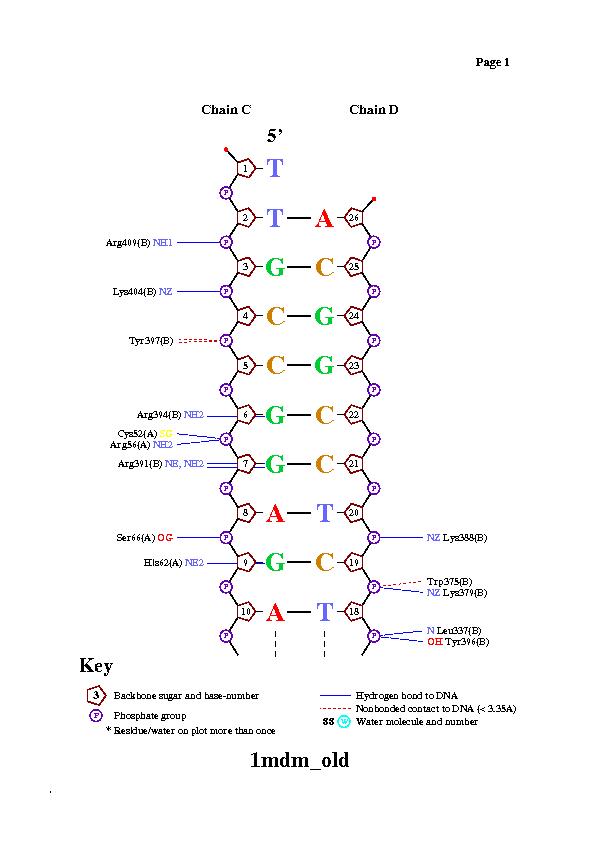
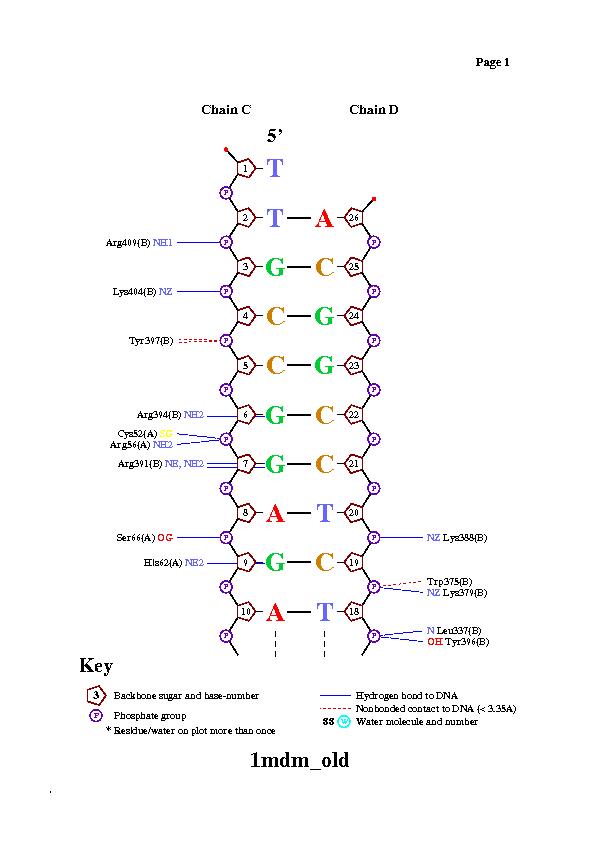
У Arg137 и Asn21 цепи А наибольшее число контактов с ДНК. Наиболее важный остаток для узнавания цепи ДНК, вероятно, Gly85, так как он связан с двумя остатками азотистых оснований, а не с остовом.
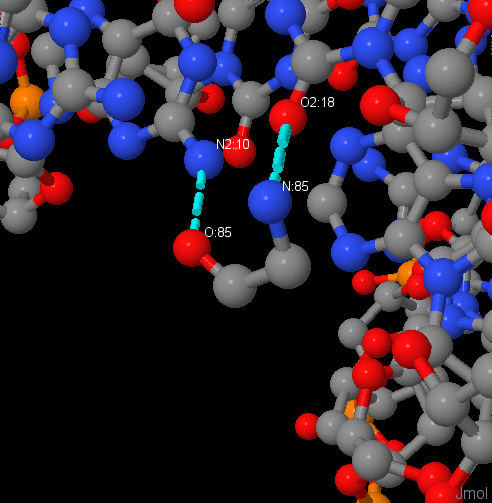
Изображение связей Gly85