И.Д.Федоров
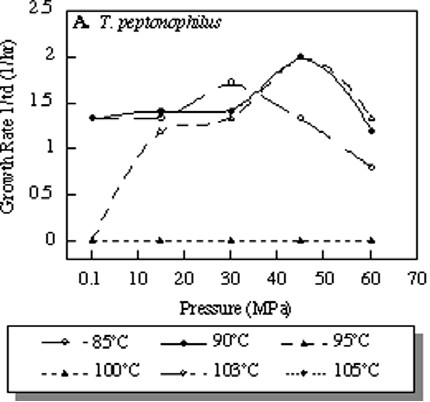
Рис. 1. Воздействие давление на рост популяции Thermococcus peptonophilus при разных температурах. Рост популяции выражен в 1/td, td – период удвоения популяции в часах. По: Kato, 2006.
Thermococcus peptonophilus – архея из семейства Thermococcaceae класса Thermococci филума Euryarchaeota. Этот вид был описан Х. Гонсалесом в 1995 году на основании образцов, полученных из гидротермальных источников в Идзу-Бонинском желобе в западной части Тихого океана (Gonzalez, 1995). Этот организм является одновременно пьезофилом и термофилом, т. е., он приспособлен к высокому давлению и высоким температурам ( Canganella, 1997).
T. peptonophilus был исследован в ряде работ. В статье Ч. Като изучены оптимальные условия обитания этого вида; установлено, что рост популяции наиболее активен при давлении около 45 MPa и температуре около 85 градусов (Kato, 2006). При этом оптимальное давление зависит от температуры: при повышении температуры оптимальное давление также повышается (рис.1). Дж. Ли было проведено исследование ДНК-полимеразы T. peptonophilus, показавшее, что у этого вида ДНК-полимераза обладает большей точностью, чем Taq-полимераза, широко применяемая для ПЦР (Lee, 2009). Исследование Х. Атоми показало, что скорость размножения T. peptonophilus достаточно высокая и достигает 1,65h-1 (Atomi, 2004). В работе Ф. Канганеллы рассматривается влияние внешних условий на выживание этого вида и показывается, что высокое давление оказывает положительное воздействие на рост популяций T. peptonophilus (Canganella, 1997).
Целью этой работы было исследовать геном T. peptonophilus и проверить, есть ли закономерности в пересечении генов на +-цепи ДНК, а также провести статистическое исследование длин белков этого вида.
Для исследования был использован геном T. peptonophilus, взятый из библиотеки NCBI. Были построены гистограммы длин пересечений с шагом десять нуклеотидов и длин белков с шагом 100 аминокислот. Кроме того, была проанализирована частота, с которой гены пересекаются. Графики были построены с помощью Google-таблиц. Для проведения теста Шапиро-Уилка была использована среда R.
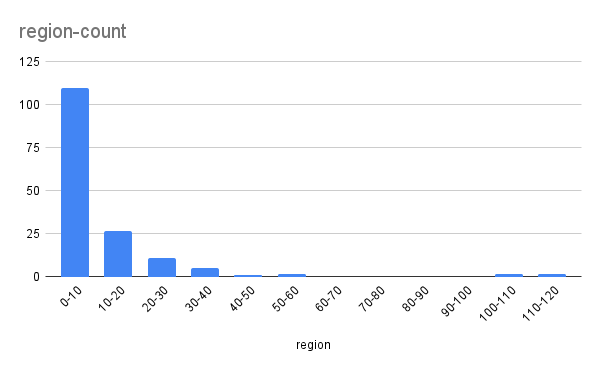
Рис. 2. Частота перекрываний разной длины. По оси X – длина перекрывания, по оси Y - частота.
Из гистограммы видно, что длина пересечений между генами в большинстве случаев меньше десяти нуклеотидов (рис. 2). Только в четырех случаях она превышает 60, достигая более 100 нуклеотидов. Максимальное пересечение – длиной в 114 нуклеотидов – между геном одного из доменов нуклеотидилтрансферазы и геном неизученного белка, а также между генами ингибитора инициации трансляции и компонента рибонуклеазы P. Средняя длина пересечения – 11 нуклеотидов; медианная – 3.
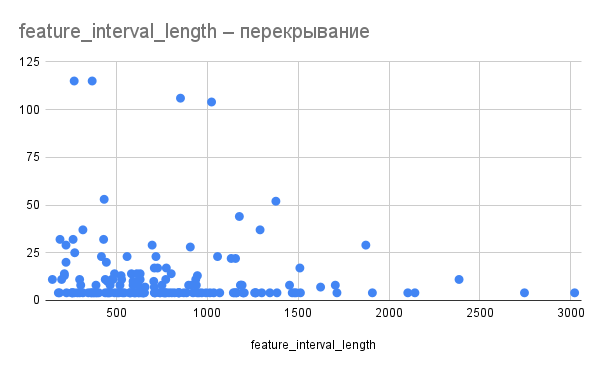
Рис. 3. Отношение длины пересечения к длине пересекающихся генов. По оси X – длина гена, по оси Y –длина пересечений со следующим геном.
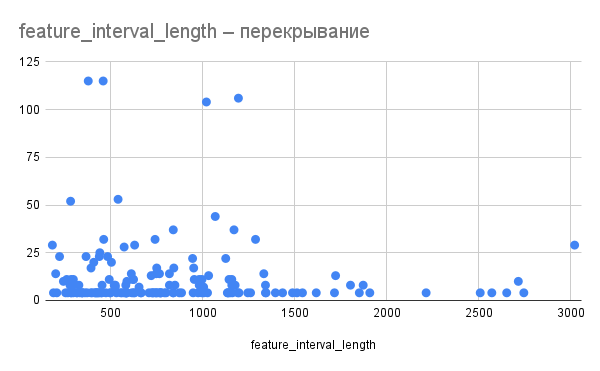
Рис. 4. Отношение длины пересечения к длине пересекающихся генов. По оси X – длина гена, по оси Y –длина пересечений с предыдущим геном.
Длина пересечения по всей видимости не зависит от длины пересекающихся генов (рис. 3, 4). Так, например, гены длиной 366 и 459 нуклеотидов перекрываются на 114, а гены длиной 1023 и 1020 – на 105.
Также можно предположить, что сильнее всего могут пересекаться гены со схожей функцией. Так, в одном из четырех случаев наиболее сильного пересечения между генами (длиной 105 нуклеотидов) в нем участвуют два схожих гена АМФ-связывающих белков. Пересечение между генами ингибитора инициации трансляции и компонента рибонуклеазы P также можно отнести к этой категории. Однако для подтверждения этой гипотезы требуется больший объем данных.
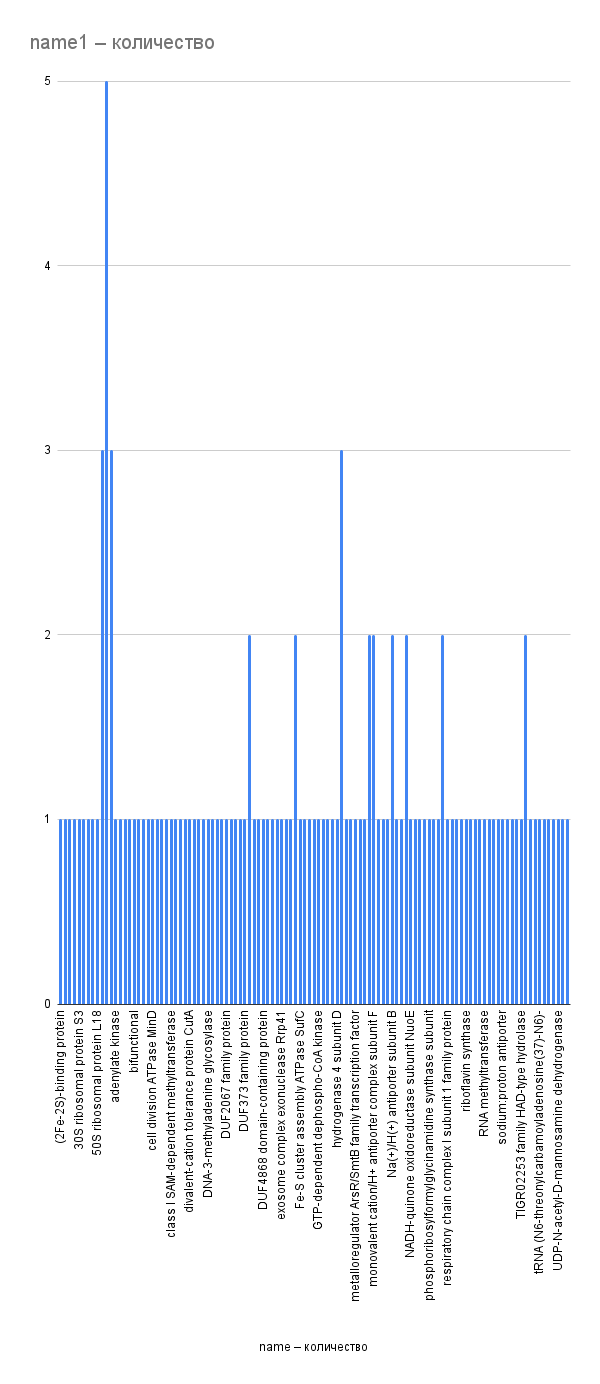
Рис. 5. Частота пересечений генов со следующими. По оси X – названия продуктов экспрессии генов, по оси Y – частота пересечений со следующим геном. Не учтены пересечения генов известных белков с генами неизвестных белков (30 раз).
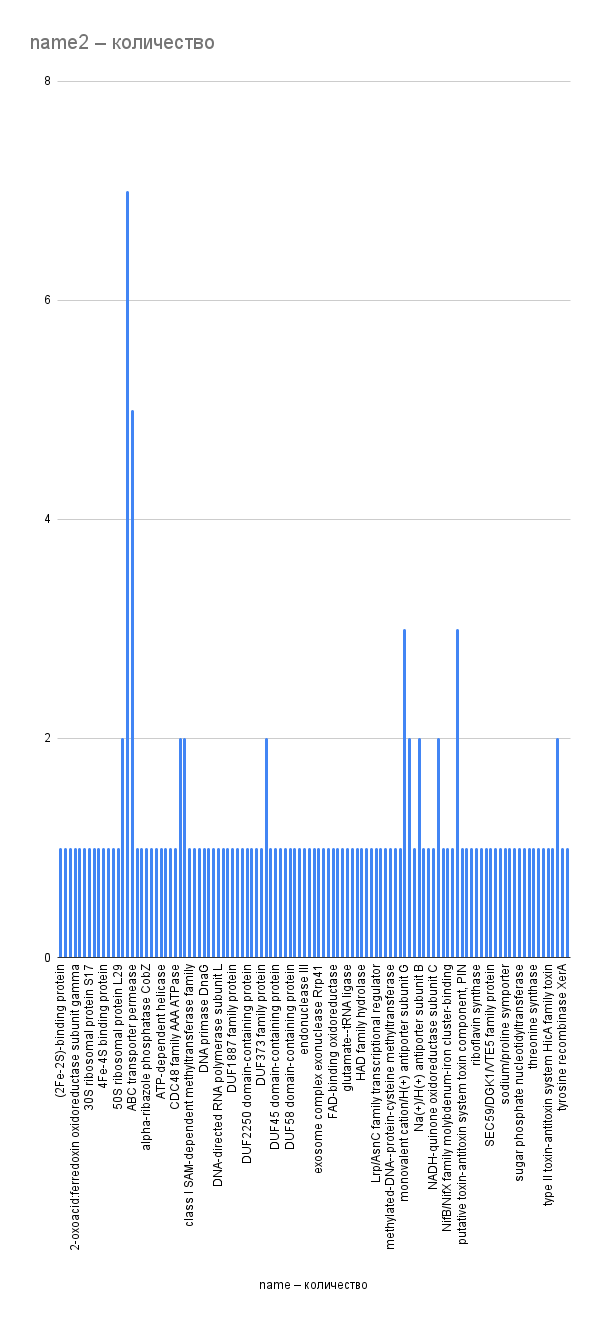
Рис. 6. Частота пересечений генов с предыдущими. По оси X – названия продуктов экспрессии генов, по оси Y – частота пересечений с предыдущим геном. Не учтены пересечения генов известных белков с генами неизвестных белков (30 раз).
Из гистограмм видно (рис, 5, 6), что большинство рассматриваемых генов пересекается с другими один-два раза; исключение составляют несколько генов, связанных с транспортными белками из семейства ABC. Ген АТФ-связывающего транспортного белка семейства ABC 12 раз участвует в пересечениях, из них семь раз – с генами ABC-пермеаз (в т. ч. пермеаз, переносящих железо). Гены ABC-пермеаз участвуют в 11 пересечениях, в т. ч. в одном случае друг с другом.
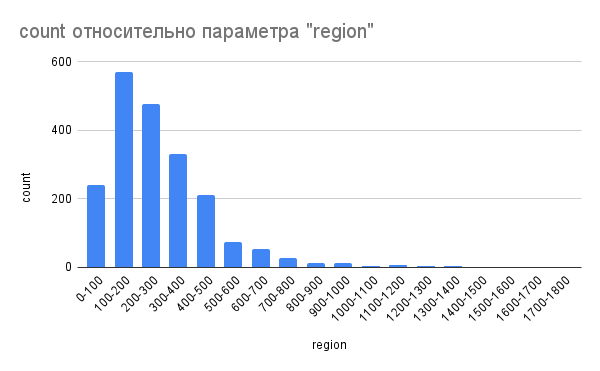
Рис. 7. Гистограмма длины белков. По оси X – длина, по оси Y – частота.
Из диаграммы (рис. 7) видно, что большинство белков T. peptonophilus имеют длину от 100 до 300 аминокислот; минимальная длина – 41, максимальная – 1746. Средняя длина – 281, медианная – 242 аминокислоты. Тест Шапиро-Уилка показал, что распределение длин белков нормальное (p-value < 2.2e-16).
Из белков с установленными функциями наименьшая длина (47 аминокислот) у доменов цинковых пальцев из группы C2H2. Эти домены как правило принадлежат белкам-факторам транскрипции и имеют небольшие размеры (Bouhouche, 2000). Таким образом, результаты анализа в этом вопросе соответствуют данным для других видов. Наибольшая длина (1746 аминокислот) у B12-зависимой редуктазы рибонуклеотидов - белка, отвечающего за синтез дезоксирибонуклеотидов из рибонуклеотидов.
Atomi,H., Fukui,T., Kanai,T., Morikawa,M., Imanaka,T. (2004). Description of Thermococcus kodakaraensis sp. nov., a well studied hyperthermophilic archaeon previously reported as Pyrococcus sp. KOD1. Archaea, 1(4), 263-267.
Bouhouche,N., Syvanen,M., Kado,C.I. (2000). The origin of prokaryotic C2H2 zinc finger regulators. Trends in microbiology, 8(2), 77-81.
Canganella,F., Gonzalez,J.M., Yanagibayashi,M., Kato,C., & Horikoshi,K. (1997). Pressure and temperature effects on growth and viability of the hyperthermophilic archaeon Thermococcus peptonophilus. Archives of microbiology, 168, 1-7.
González,J., Kato,C. Horikoshi,K. (1995). Thermococcus peptonophilus sp. nov., a fast-growing, extremely thermophilic archaebacterium isolated from deep-sea hydrothermal vents. Arch Microbiol, 164, 159–164.
Kato,C. (2006). 31 Handling of Piezophilic Microorganisms. In Methods in microbiology (Vol. 35, pp. 733-741). Academic Press.
Lee,J.I., Kim,Y.J., Bae,H., Cho,S.S., Lee,J.H., Kwon,S.T. (2010). Biochemical properties and PCR performance of a family B DNA polymerase from hyperthermophilic Euryarchaeon Thermococcus peptonophilus. Applied biochemistry and biotechnology, 160, 1585-1599.