Практикум 12.
Задание 1. Построить выравнивание представителей домена Pfam белков с разной доменной архитектурой
Описание выбранного домена из Pfam:
AC Lig_chan
ID PF00060
Функция домена - лиганд-зависимый ионный канал.
Ссылка на страницу домена в Pfam http://pfam.xfam.org/family/PF00060#tabview=tab0
Список доменных архитектур с выбранным доменом представлен на странице http://pfam.xfam.org/family/PF00060#tabview=tab1
Описание выбранных доменных архитектур
Архитектура 1: 1282 последовательности

Домены ANF_receptor, Lig_chan-Glu_bd, Lig_chan
ANF_receptor (PF01094) - внеклеточные лиганд-связывающие домены разных рецепторов.
Lig_chan-Glu_bd (PF10613) - Ligated ion channel L-glutamate- and glycine-binding site.
Домен, расположенный в просвете канала upstream от первого трансмембранного домена трансмембранных ионных каналов.
Связывает L-глутамат и глицин. Ассоциирован с доменом Lig_chan (PF00060).
Архитектура 2: 139 последовательностей

Домены ANF_receptor, Lig_chan-Glu_bd, Lig_chan, NMDAR2_C
NMDAR2_C (PF10565) - N-methyl D-aspartate receptor 2B3 C-terminus. Этот домен находится на С-конце многих NMDA-рецепторов, частов стречается в доменных архитектурах, содержащих Lig_chan upstream и ANF_receptor.Высоко гидрофилен. Предсказано, что он является крупным внеклеточным доменом. Содержит несколько потенциальных сайтов N-гликозилирования.
Описание выбранного таксона
В качестве таксона выбран Euteleostomi ("костные позвоночные", таксон включает костных рыб и тетрапод).
В качестве подтаксонов были выбраны: Mammalia (млекопитающие)
и Actinopterygii (лучеперые рыбы).
Результаты работы:
Ссылка на таблицу Excel с описанием всех белков из Uniprot, включающих домен (лист "все"), и выборкой представителей (лист "выбранные").
Выравнивание отобранных последовательностей домена, разбитое на группы по доменным архитектурам и раскрашенное по консервативности внутри групп.
В качестве порога окраски выбрано 40%. В выравнивани ясно видны домены - в виде протяженных блоков с высокой консервативностью колонок. Отсутствуют гэпы в консервативных колонках. В процессе редактирования было удалено около 80 а.к. с N-конеца, поскольку там не было выравнивания (отсутствовали консервативные колонки, крайне много гэпов. На С-конце выравнивания для доменной архитектуры 2 присутствует хорошее выравнивание, поэтому я его оставила.
Задание 2: построение филогенетического дерева домена
Я удалила из выравнивания колонки, не относящиеся к выбранному домену (определила его границы исходя их данных о доменной архитектуры). Добавила к началу последовательностей мнемоники архитектур (1 и 2) и подтаксонов (A - Actinopterygii, M - Mammalia). (Например, 1_A -архитектура 1, подтаксон Actinopterygii).
Затем было построено дерево программой MEGA методом NJ с параметрами: 100 бутстрэп реплик
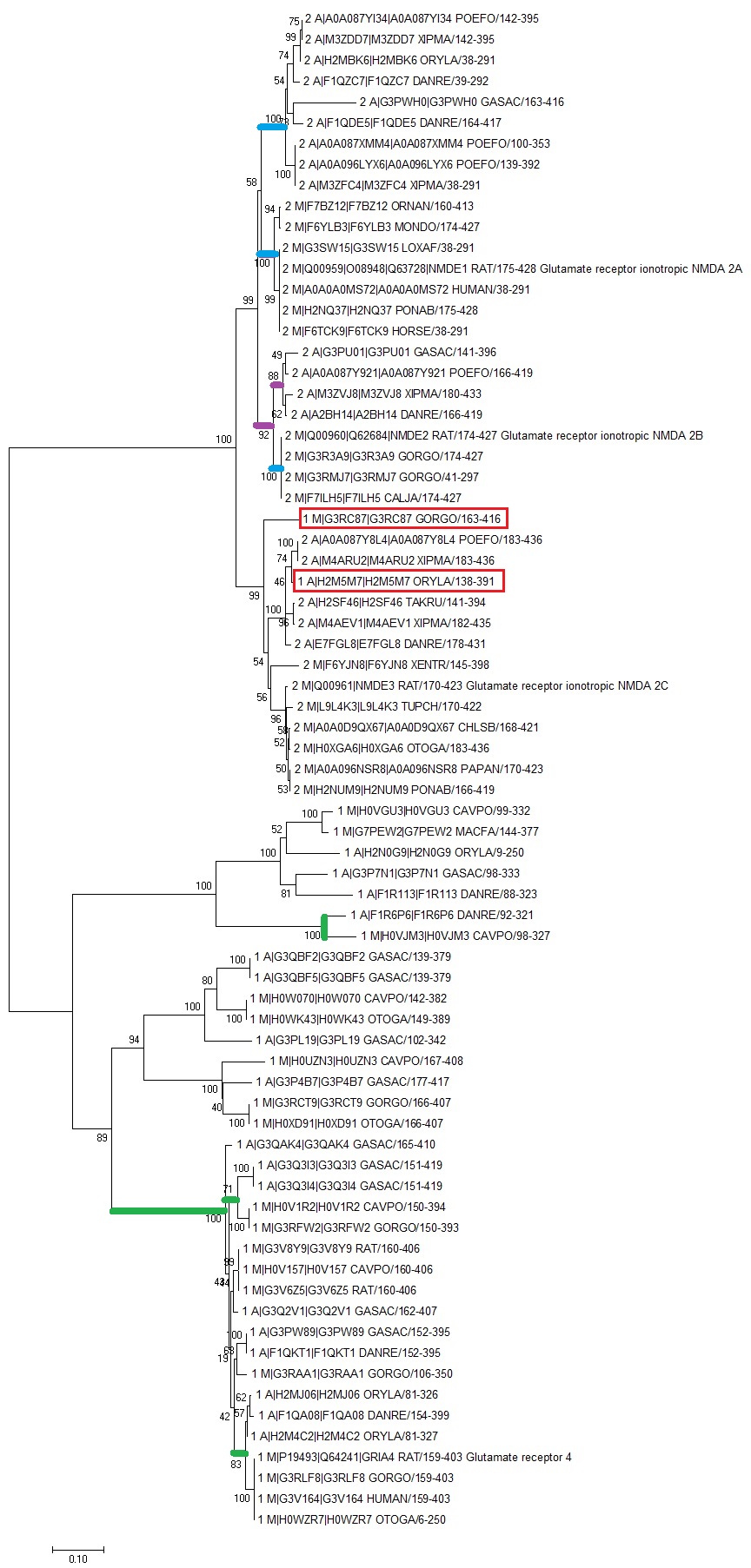
Рис.3 Дерево, полученное по выравниванию домена из доменных архитектур 1 и 2. Красным отмечены последовательности белков, в которых происходила доменная перестройка.
Файл со скобочной формулой дерева в формае newvick
На полученном дереве последовательности, соответствующие разным доменным архитектурам, разделились на две соответствующие ветви, за исключением двух случаев (отмечены красным на рис.3). Это свидетельствует о том, что данные белки разошлись после того, как новый домен (NMDAR2_C, который есть в структуре 2, но отсутствует в структуре 1), был приобретен. То есть приобретение домена NMDAR2_C было давно произошедшим однократным событием. Два белка с доменной архитектурой 1 (1_M_G3RC87 И 1_M_H2M2M7) попали в ветвь второй доменной архитектуры. Это свидетельствует о том, что они имели NMDAR2_C домен и потеряли его относительно недавно.
Внутри некоторыхболее мелких ветвей наблюдается разбиение на ветви по подтаксонам (например, синие, фиолетовые ветви). Это означает, что рассматриваемые домены этих белков у представителей разных подтаксонов расходились от общего белкового предка позже, чем разошлись от общего предка эти таксоны. В целом, это не очень удивительно, поскольку разнообразие среди лиганд-зависимых ионных каналов довольно велико и, по-видимому, их строение таксоноспецифично.
Некоторые ветви объединяют последовательности из разных подтаксонов (например, выделенные зеленым). Это означает, что у общего предка двух подтаксонов уже был данный домен, отделившийся от предкового домена. Таких ветвей довольно много.