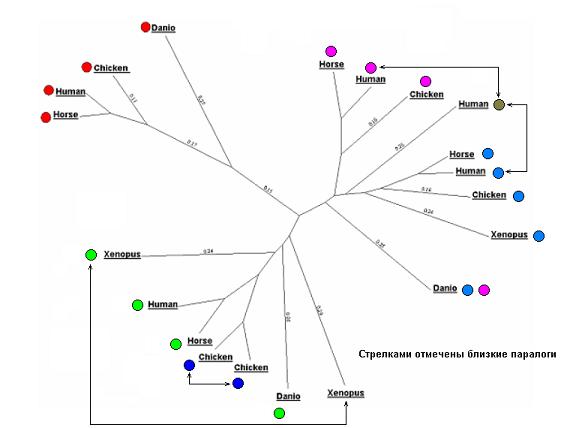
Анализ “правой” ветви.
Сверху расположены три последовательности, отмеченные фиолетовым, точно являющиеся ортологами. Ниже находятся еще 4 последовательности, отмеченные голубым и тоже точно являющиеся ортологами. На ортологичность последовательностей в этих группах указывает соответствие их взаимного расположения внутри групп известному ходу эволюции животных.
Последовательность рыбки данио равновероятно является ортологом к последовательностям одной из этих групп. Возможно, если вторая последовательность из данио (ортолог к оставшейся из групп) не просто не включена в дерево, а вообще не существует в природе, удвоение гена произошло на стадии между рыбами и земноводными с последующим образованием двух паралогичных групп последовательностей. Точные ортологические отношения для последовательности из данио не разрешимы.
Последовательность из человека, помеченная коричневым, очевидно, является паралогом к двум другим последовательностям из человека в группах снизу и сверху. Эволюционно, видимо, в правой ветви на стадии между рыбами и земноводными произошло образование трех паралогичных генов (конечно, в два этапа), которые потом дали три группы генов в представителях более эволюционно высоких классов. Последовательности из ксенопуса, цыпленка и лошади, ортологичные «коричневой» последовательности из человека, либо просто не включены в дерево, либо вообще эволюционно не сохранились.
Анализ “левой” ветви.
Последовательности, отмеченные зеленым, и одна из «синих» последовательностей из цыпленка, являются ортологами. Какая конкретно последовательность из цыпленка, установить нельзя. Сами эти две последовательности из цыпленка являются между собой паралогами.
Последовательность из ксенопуса, никак не отмеченная, является паралогом к последовательности из ксенопуса, отмеченной зеленым. Восстановить правдоподобный ход эволюционных событий для этой («бесцветной») последовательности не представляется возможным.