
<< BACK
Для сравнения глобального и локального парных выравниваний были взяты два белка - DNAK_HALMA и HSP7C_HUMAN. Выравнивания были построены с помощью команд пакета EMBOSS. ЕMBOSS содержит две программы: needle - для глобального выравнивания и water - для локального выравнивания. Затем программа EMBOSS вычисляет основные свойства выравниваний,результат которых представлен в таблице 1. Для обоих программ штраф за гэпы следующие: gapopen 10,0, gapextend 0,5; для глобального выравнивания endweight Y, endopen 10,0, endextend. Матрица весов для обеих программ - EBLOSUM62. Локальное и глобальное выравнивания отличаются концевым участком: в локальном выравнивании последовательности обрезаны по консервативным 612 и 639 остаткам глицина, так как за ними почти нет участков гомологии и присутствуют большие индели. За счет этого локальное выравнивание становится короче - вырезается два инделя. В остальных позициях оба выравнивания идентичны, но вырезание лишних гэпов дает локальному выравниванию больший вес. В этом случае локальное выравнивание более информативно при меньших размерах, поскольку чаще приходится сравнивать внутрибелковые структуры, а не концевые, так как первые больше подверженны эволюционным факторам.
| Тип выравнивания | Длина выравнивания | Число гэпов | Процент гэпов | Число инделей | а.к.п | а.к.п, % | ф.к.п | ф.к.п, % | Вес выравнивания |
| Локальное (water) | 650 | 49 | 7,54 | 15 | 302 | 46,46 | 416 | 64,00 | 1378 |
| Глобальное (needle) | 673 | 65 | 9,66 | 17 | 305 | 45,32 | 420 | 62,41 | 1367 |
Рис 1. Глобальное выравнивание DNAK_HALMA и HSP7C_HUMAN. Окрашено по схеме ClustalX, 100% идентичность.

Рис 2. Локальное выравнивание DNAK_HALMA и HSP7C_HUMAN. Окрашено по схеме ClustalX, 100% идентичность.
Таким образом, локальное и глобальное выравнивания представляют собой в определенной степени идентичные выравнивания, отличающиеся концевыми участками. Глобальное показывает гомологию между концевыми остатками, тогда как в локальном наоборот.
Для анализа выравниваний негомологичных белков были построены локальные выравнивания аминокислотной последовательности белка MarR (ADX57967.1) попарно с последовательностями белка ANE87960.1 (белок репарации алкилированной ДНК), ацетил - КоА синтазой (BAC69532.1), гликозил - трансферазой (ADX55361.1), AKQ41279.1, AMD46139.1. При построении выравнивания мной использовалась программа water EMBOSS, штраф за индель 10.0, за каждый следующий гэп 0.5, матрица весов - EBLOSUM62.
| GenBank | Длина последовательности | Длина выравнивания | Число гэпов | Процент гэпов | Число инделей | а.к.п | а.к.п, % | ф.к.п | ф.к.п, % | Вес выравнивания |
| AAV48029.1 | 635 | 650 | 38 | 5,85 | 11 | 302 | 46,46 | 416 | 64,00 | 1378 |
| AAH19816.1 | 646 | 650 | 11 | 1,69 | 4 | 302 | 46,46 | 416 | 64,00 | |
| ADX57967.1 | 166 | 50 | 0 | 0,00 | 0 | 15 | 30,00 | 24 | 48,00 | 36,5 |
| ANE87960.1 | 237 | 50 | 3 | 6,00 | 2 | 15 | 30,00 | 24 | 48,00 | |
| ADX57967.1 | 166 | 127 | 19 | 14,96 | 4 | 31 | 24,41 | 52 | 40,94 | 43,5 |
| BAC69532.1 | 715 | 127 | 13 | 10,24 | 3 | 31 | 24,41 | 52 | 40,94 | |
| ADX57967.1 | 166 | 154 | 16 | 10,39 | 4 | 38 | 24,68 | 54 | 35,06 | 58,0 |
| ADX55361.1 | 439 | 154 | 28 | 18,18 | 2 | 38 | 24,68 | 54 | 35,06 | |
| ADX57967.1 | 166 | 174 | 31 | 17,82 | 6 | 40 | 22,99 | 64 | 36,78 | 44,0 |
| AKQ41279.1 | 768 | 174 | 7 | 4,02 | 2 | 40 | 22,99 | 64 | 36,78 | |
| ADX57967.1 | 166 | 208 | 45 | 21,63 | 6 | 42 | 20,19 | 73 | 35,10 | 45,5 |
| AMD46139.1 | 452 | 208 | 20 | 9,62 | 4 | 42 | 20,19 | 73 | 35,10 |
Процент гэпов в локальном выравнивании негомологичных последовательностей ДОЛЖЕН БЫТЬ значительно больше, а процент абсолютно консервативных и функционально консервативных позиций - меньше. На рисунках 3 и 4 представлены выравнивания негомологичных последовательностей ADX57967.1 и ANE87960.1; ADX57967.1 и AMD46139.1, соответственно. Первое выравнивание в несколько раз короче самих последовательностей и представляет собой концевые участки, в которых обнаружилось НАИБОЛЬШЕЕ сходство. Его (выравнивания) параметры консервативности имеют значения несколько больше, чем у некоторых остальных выравниваний ПРЕДПОЛОЖИТЕЛЬНО негомологичных последовательностей, и в примерно на столько же меньше, чем соответствующие параметры у выравнивания гомологичных последовательностей (например, 1-ая пара в табл.). Мной было построено глобальное выравнивание ADX57967.1 и ANE87960.1 (Рис. 5) и выяснено, что локальное выравнивание ПОЛНОСТЬЮ СООТВЕТСТВУЕТ части глобального. Это натолкнуло меня на мысль, что эти участки ВЕРОЯТНО действительно гомологичны, учитывая, что ОБА БЕЛКА МОГУТ ВЗАИМОДЕЙСТВОВАТЬ С ДНК (т.е. функционально схожи). На рисунке 5 я выделил этот участок, который, по моему мнению, можно рассмотреть как болк. Выравнивание последней (относительно табл.) пары последовательностей (рис.4) представляет собой чередование в шахматном порядке участков с гэпами и а.к.о. А это точно НЕ свидетельствует о гомологии данных последовательностей.

Рис.3 Выравнивания негомологичных последовательностей ADX57967.1 и ANE87960.1.
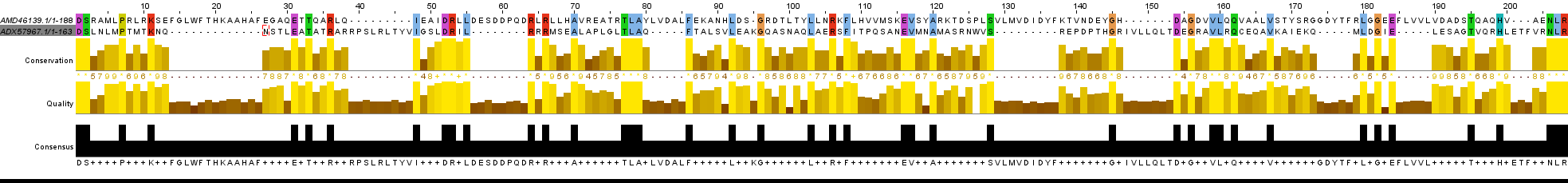
Рис.4 Выравнивания негомологичных последовательностей ADX57967.1 и AMD46139.1

Рис.5 Глобальное выравнивание последовательностей ADX57967.1 и ANE87960.1.
Используемая цветовая схема позволяет оценить консервативность выравнивания. Предполагается, что применяемая цветовая схема будет окрашивать функционально идентичные остатки в бледно-голубой, отчасти это имеет место. Таким образом, мы можем продемонстриовать наиболее консервативные участки. Можно с уверенностью утверждать, что выравнивающие программы предоставляют возможность отличать гомологичные и не гомологичные белки на основе описанных признаков.
Для сравнения парных выравниваний, полуженных из множественного выравнивания Jalview, программами needle и water из EMBOSS я взял два белка использованных ранее - DNAK_HALMA и HSP7C_HUMAN. В программе Jalview я сравнил три выравнивания гомологичных последовательностей в одном окне, назначив каждому свою группу. Выравнив выравнивания относительно друг друга, не меняя колонки внутри выравниваний, я нашел некоторые отличия. Как уже было отмечено выше, локальное и глобальное выравнивания отличаются только концевыми участками, так что речь пойдет об отличиях этих парных выравниваний от части множественного выравнивания. Верхние две строчки - глобальное выравнивание, две посередине - локальное, две снизу - часть от множественного выравнивания.

Рис.5 Глобальное выравнивание последовательностей ADX57967.1 и ANE87960.1.
Становится возможным сделать вывод: все выравнивания в общем-то идентичны, однако есть несколько несоответствий. В позиции 78 фенилаланин в верхней последовательности стоит в колонке с гэпом в нижней в локальном и глобальном выравниваниях. В множественном в этой позиции стоит гэп в верней и аспарагиновая кислота в нижней. Фенилаланин в множественном выравнивании стоит в 79 позиции напротив тирозина, последний в других выравниваниях стоит в 104 позиции. В позиции 113 глутаминовая кислота стоит напротив лизина во множественном выравнивании, однако в парных выравниваниях этот а.к.о. стоит в 111 позиции напротив так же глутаминовой кислоты. Группа остатков IDSLY в позициях выравнивания 288 - 292 стоит напротив группы IATTD в парных выравниваниях, во множественном она сдвинута влево на три позиции и гомологична LPFIA. Глутаминовая кислота и глицин из позиций 294 - 295 множественного выравнивания гомологичны глицину и пролину, в то время как в парных выравниваниях они стоят в позицях 293 - 294 и гомологичны аспарагиновой кислоте и глицину, образуют косервативность в позиции 294 по глицину. Наиболее правдоподобное выравнивание - это парное локальное. Но имеются некоторые проблемы, связанные с гомологичностью внутренних остатков (гомологичность парных остатков, находящихся "под вопросом", представляется более вероятной) и ведет себя странно в конце последовательностей. Сравнивая локальное и глобальное выравнивания, нужно сказать, что в этом конкретном случае нет необходимости в выравнивании конечного «хвоста», так как с точностью нельзя предположить, что произошло в ходе эволюции.