Комплексы ДНК-белок





Для тРНК из PDB-структуры 1H4S я сравнила реальную вторичную структуру с предсказанными программами einverted и RNAfold (алгоритм Зукера). С помощью программы einverted (на вход принимает последовательность нуклеотидов, на выходе 2 файла - *.inv (комплементарные участки) и *.fasta (водородные связи)) из пакета EMBOSS я искала инвертированные участки в нуклеотидных последовательностях. При стандартных значениях (gap penalty [12], minimum score threshold [50], match score [3], mismatch [-4]) программа выдает пустой файл. При уменьшении minumum score threshold до 30 также получался пустой файл. При уменьшении minimum score threshold до 10 было найдено 7 пар.
SEQUENCE: Score 21: 7/7 (100%) matches, 0 gaps
1 cggggag 7
|||||||
73 gcccctc 67
Затем мне пришлось уменьшить minimum score threshold до 0, а также изменить некоторые другие параметры. Результат при значениях gap penalty [12], minimum score threshold [0], match score [2], mismatch [-1]:
SEQUENCE: Score 23: 19/34 ( 55%) matches, 0 gaps
2 ggggagtagcgcagcccggtagcgcacctcgttc 35
|| || | | | | | || | |||||| |
70 cctctgacctagacttggtcgcggggggagcagg 37
Как можно заметить, здесь достаточно много гэпов, так что достоверность такого файла не особенно велика. Поиск стеблей не очень эффективен, так как данная программа учитывает только канонические взаимодействия (неканонических все же достаточно много).
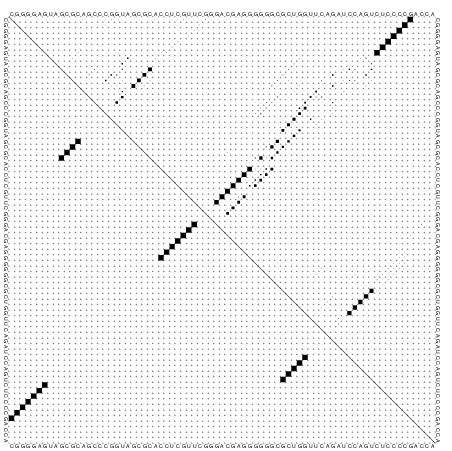
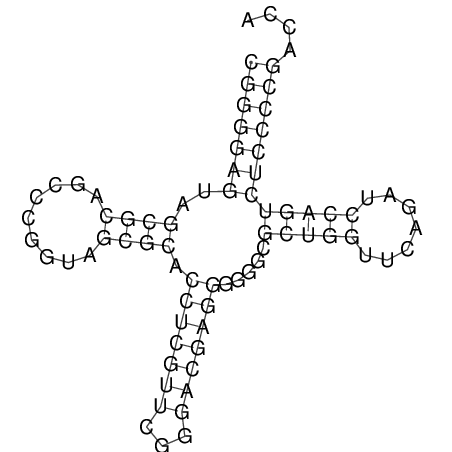
Затем была использована программа RNAfold из пакета Vienna Rna Package, которая реализует алгоритм Зукера. В результате была получена вторичная структура тРНК, изображенная на рисунке 2, на рисунке 1 изображена полученная матрица. Структура, полученная со стандартными параметрами программы, имеет минимальную свободную энергию -31.50 kcal/mol, представленная структура, наиболее похожая на нормальную тРНК, имеет энергию -26.49 kcal/mol.
В таблице 1 произведено сравнение вторичных структур тРНК, полученных с помощью программ find_pair, einverted и RNAfold. Указано количество пар нуклеотидов в стеблях тРНК по результатам трех программ.
| Участок структуры | Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
| Акцепторный стебель | 5'-4-7-3' 5'-66-69-3' 4 пары |
предсказано 0 пар |
предсказано 0 пар (смещение на одно основание) |
| D-стебель | 5'-10-13-3' 5'-22-25-3' 4 пары |
предсказано 0 пар |
предсказано 0 пар (смещение на одно основание) |
| T-стебель | 5'-49-53-3' 5'-61-65-3' 5 пар |
предсказано 0 пар |
предсказано 0 пар (смещение на одно основание) |
| Антикодоновый стебель | 5'-38-44-3' 5'-26-32-3' 7 пар |
предсказано 0 пар |
предсказано 7 пар (все) |
| Общее число канонических пар нуклеотидов | 23 |
19 |
21 |
Далее я работала с ДНК-белковым комплексом из PDB-структуры 1MHD. Для начала я определила следующие группы атомов ДНК: множество атомов кислорода 2'-дезоксирибозы (*.O?'); множество атомов кислорода в остатке фосфорной кислоты (*.OP?) и множество атомов азота в азотистых основаниях (*.N?). Эти множества были названы в JMol set1, set2 и set3 соответственно. В данном скрипте определяются эти группы атомов. Также я написала скрипт, вызов которого в JMol последовательно показывает всю структуру; только ДНК в проволочной модели; ДНК с выделенными шариками множеством атомов set1; set2 и set3. Окраска ДНК производится по cpk. В приведенном апплете можно увидеть все изображения.
В таблице 2 представлена информация о контактах разного типа (полярных и неполярных) между ДНК и белком в структуре 1MHD. Полярными атомами считаются атомы кислорода и азота, неполярными - атомы углерода, фосфора и серы. Полярный контакт - ситуация, в которой расстояние между полярным атомом белка и полярным атомом ДНК меньше 3.5Å. Неполярный контакт - пара неполярных атомов на расстоянии меньше 4.5Å.
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 1 |
9 |
10 |
| остатками фосфорной кислоты | 8 |
0 |
8 |
| остатками азотистых оснований со стороны большой бороздки | 2 |
9 |
11 |
| остатками азотистых оснований со стороны малой бороздки | 0 |
3 |
3 |
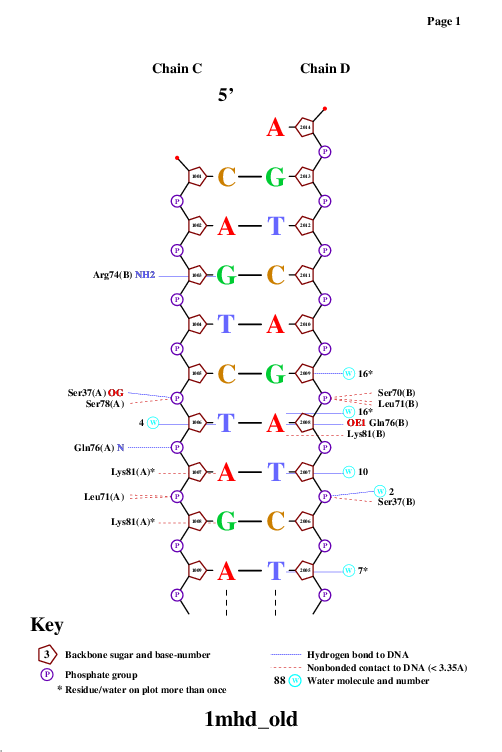
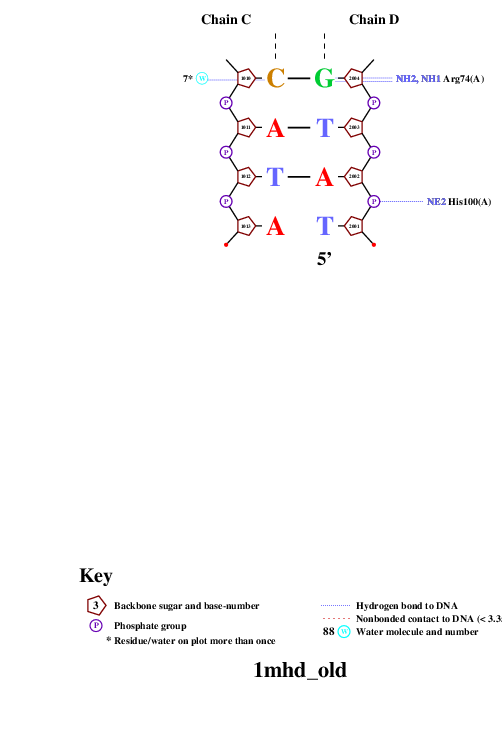
Затем я попыталась оценить ДНК-белковые контакты с помощью программы nucplot, используя файл в старом формате.
Белок образует достаточно много контактов с сахаро-фосфатным остовом ДНК. Скорее всего, эти контакты нужны для того, чтобы белок удерживался на ДНК. Однако контактов с остатками азотистых оснований также достаточно много. Эти контакты отвечают за распознавание белком специфической последовательности нуклеиновой кислоты, что является важным в данном случае, так как данный белок принимает участие в одном из процессов биосигналинга (TGF-beta signaling).Также можно сказать, что неполярные контакты встречаются чаще, чем полярные, но, скорее всего, именно полярные контакты отвечают за специфическое распознование.
Аминокислотный остаток с наибольшим числом указанных на схеме контактов с ДНК - Arg74, ибо он образует контакты с NH2 в двух местах и c NH1 - в одном.
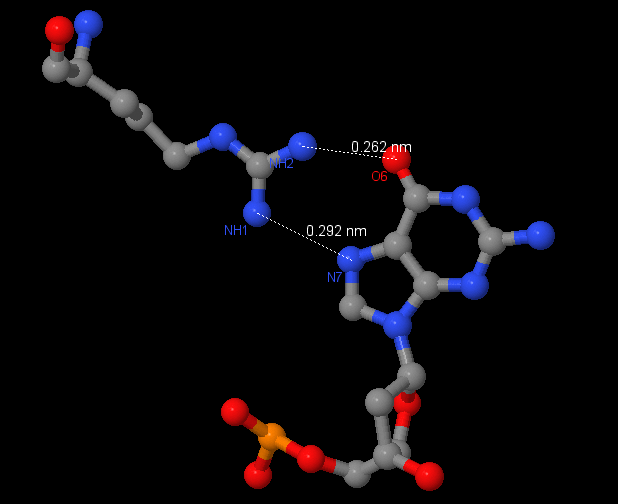
Рис.3.Контакты Arg74 и G2004.
Аминокислотный остаток, наиболее важный для распознавания последовательности ДНК - Gln76 (связывается с самим азотистым основанием, хорошо ложится в бороздку ДНК).
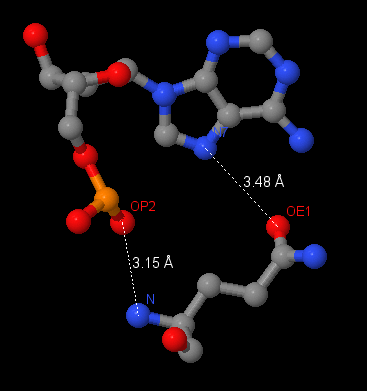
Рис.4.1.Контакты Gln76 с A2008.
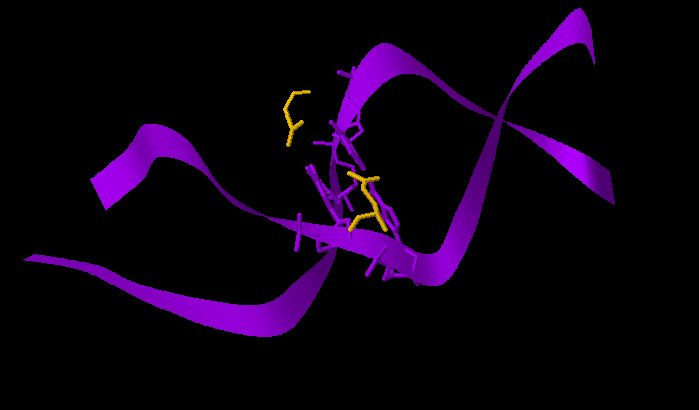
Рис.4.2. Gln76 в бороздке ДНК (глутамин отмечен желтым)
Ссылки:
[1] О. О. Фаворова. Строение транспортных РНК и их функция на первом (предрибосомном)
этапе биосинтеза белков. Соровский образовательный журнал, №11 (1998): 71-77, рис.3.