Контакты
Ленинские горы МГУ 1, стр. 73
8-916-939-49-78
jiabicht@rambler.ru
vk.com/allweiss
8-916-939-49-78
jiabicht@rambler.ru
vk.com/allweiss
Филогенетическое дерево бактерий было построено по выравниванию генов 16S rRNA (таблица 1).
| Организм | Мнемоника | AC EMBL | Координаты (F/R) 16S rRNA |
| Bacillus subtilis | BACSU | D26185 | 73411..74963 (F) |
| Clostridium botulinum | CLOB1 | CP000726 | 9282..10783 (F) |
| Finegoldia magna | FINM2 | AP008971 | 197837..199361 (F) |
| Geobacillus kaustophilus | GEOKA | BA000043 | 10421..11973 (F) |
| Lactobacillus delbrueckii | LACDA | CP000412 | 43705..45265 (F) |
| Lactococcus lactis | LACLM | CP001834 | 529828..531363 (F) |
| Staphylococcus epidermidis | STAES | AE015929.1 | 1598006..1599559 (R) |
| Streptococcus pneumoniae | STRPN | AE005672 | - |
Таблица 1. Организмы и гены 16S rRNA, для которых было построено филогенетическое дерево.
Для огранизма Streptococcus pneumoniae ген 16S rRNA не аннотирован в геноме, поэтому его последовательность была получена
при помощи nblast против полного генома с использованием последовательности 16S rRNA из Bacillus subtilis.
Последовательности гена 16S rRNA были выровнены в программе JalView 2.8 алгоритмом Muscle (параметры по умолчанию)
(Выравнивание).
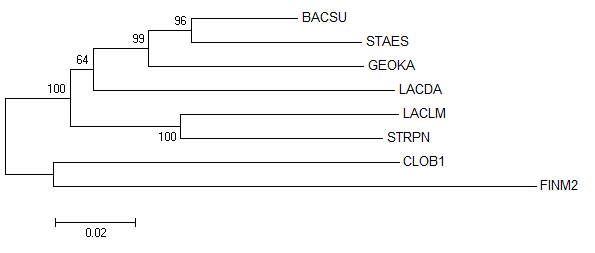
По выравниванию в программе MEGA 6.06 было построено филогенетическое дерево (Рис. 1, 2) алгоритмом Maximum Likelihood с 200 циклами
Bootstrap анализа.
| 
|
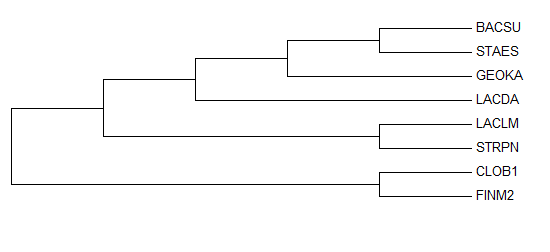
| Рис. 1. Филогенетическое дерево бактерий, построенное по гену 16S rRNA. Указаны поддержки ветвей. | Рис. 2. Филогенетическое дерево бактерий, построенное по гену 16S rRNA, на основе bootstrap анализа. |
Реконструированной дерево отличается от правильного не значительно (Рис. 3).
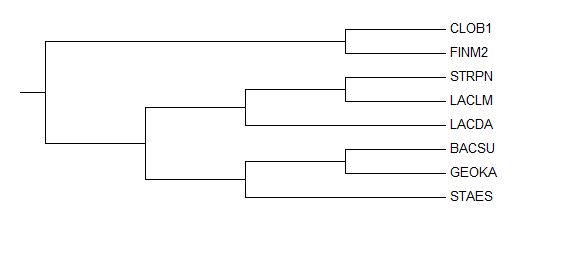
Рис. 3. Правильное филогенетическое дерево бактерий.
Как видно, на построенном на основе гена 16S rRNA филогенетическом дереве 2 неправильные ветви:
{BACSU, STAES} и {BACSU, STAES, GEOKA, LACDA}.
Два гомологичных белка будем называть ортологами, если они а) из разных организмов;
б) разделение их общего предка на линии, ведущие к ним, произошло в результате видообразования.
Два гомологичных белка из одного организма будем называть паралогами.
Для белка CLPX_BACSU из организма Bacillus subtilis при помощи blast были найдены гомологи из организмов из таблицы 1.
Все эти белки являются АТФ-зависимыми протеазами, или связывающими АТФ субьединицами протеаз.
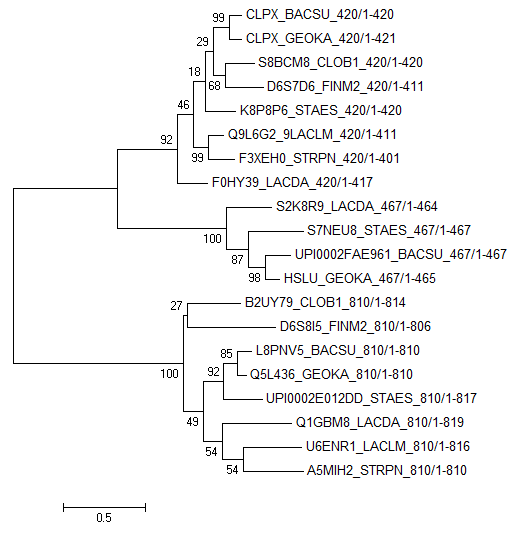
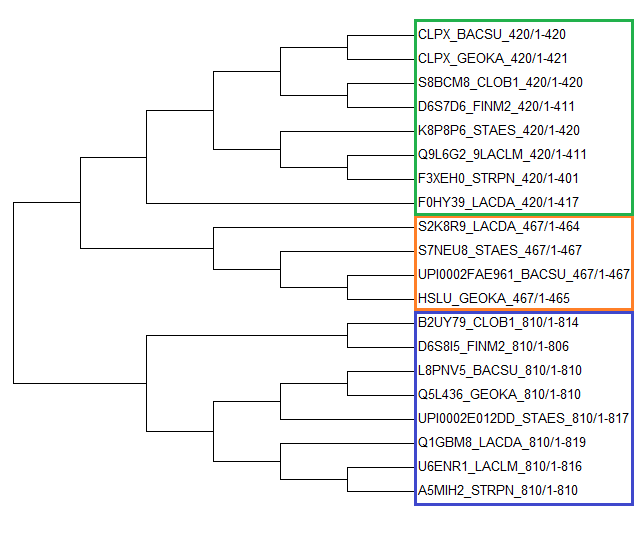
Все найденные белки группировались в 3 блока по длине:
| 
|
| Рис. 4. Филогенетическое дерево бактерий, построенное по гену белка CLPX. Указаны поддержки ветвей. | Рис. 5. Филогенетическое дерево бактерий, построенное по гену белка CLPX, на основе bootstrap анализа. Разноцветными рамками выделены блоки последовательностей разной длины. |
Как видно, по дереву на рис. 5,- последовательности с разными длинами хорошо группируются в отдельные ветви. Это подтверждает гепотезу
о том, что они являются паралогами.
Что интересно, ветвь с белками длиной 810 в точности повторяет правильное дерево бактерий (Рис. 3),
тогда как ветвь 410 содержит ошибки, хотя именно к ней относится белок CLPX_BACSU, при помощи которого были найдены все остальные белки.
По дереву можно сказать, что дупликация гена произошла еще у общего предка всех этих бактерий, поскольку варианты 810 и 410 встречаются
у всех этих бактерий. Видно, что вторая дупликация гена, приведшая к образованию варианта 467 (сложно сказать, от какого из вариантов - 810 или 410)
произощла у общего предка бактерий {BACSU, GEOKA, STAES, LACDA}. Интересно, что у бактерий LACLM и STRPN этого варианта нет, хотя судя по правильному
дереву должен бы быть.
Однако, если принять правильность дерева, построенного по гену 16S rRNA, где ветвь {LACLM, STRPN} является сестринской ветвью
к {BACSU, GEOKA, STAES, LACDA}, то все становится на свои места.
Исходя из этих независимых данных можно попробовать сделать вывод о том,
что "правильное" дерево не совсем верно, и порядок Lactobacillales (STRPN, LACLM, LACDA) является парафилетической группой.