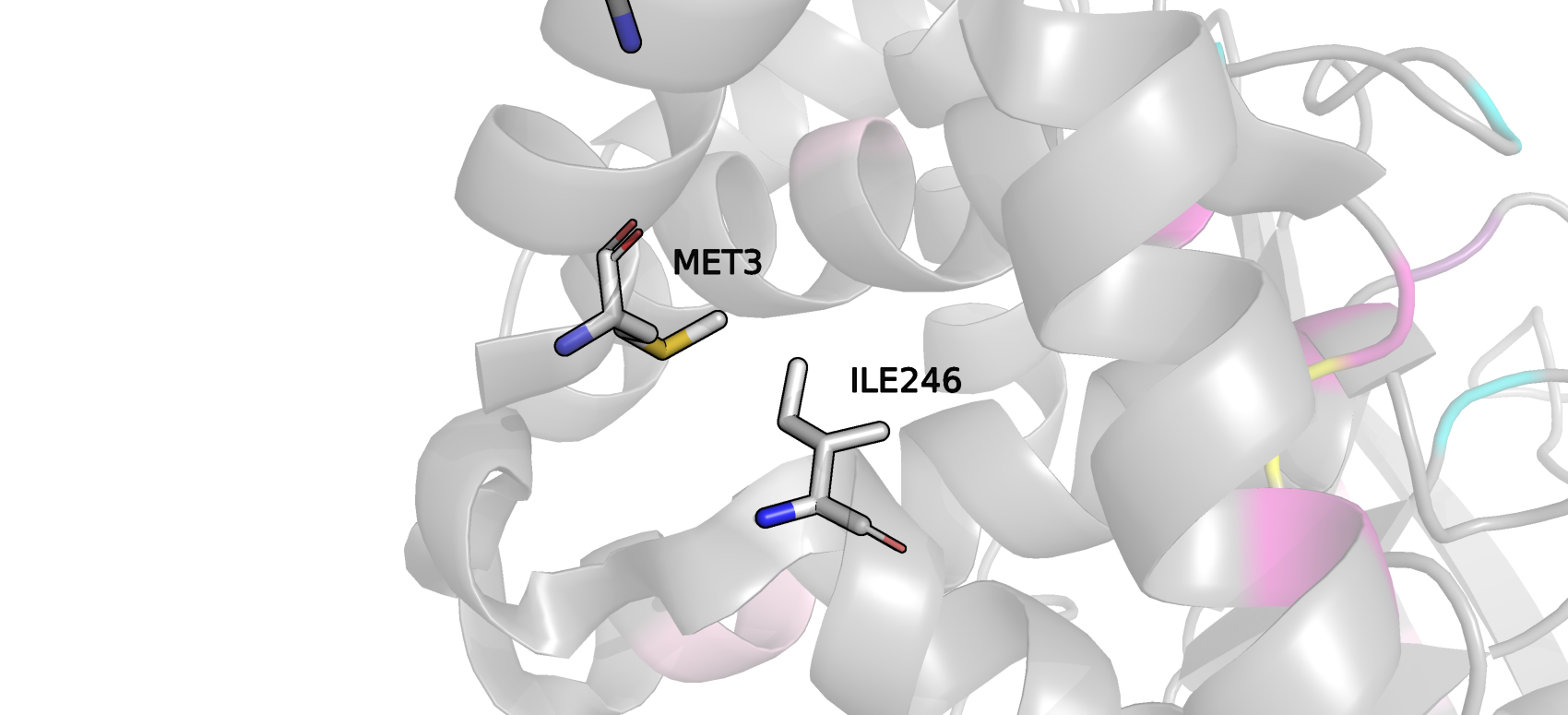Сообщим краткие сведения о белке. Его PDBid 2zmm. Белок является протеин-тирозинфосфатазой 1B (PTP-1B), предшественником внутриклеточной фосфотазы, которая участвует
в негативной регуляции путей передачи сигналов от инсулина и лептина. Он был выделен методом РСА с разрешением в 2.10 A для поиска низкомолекулярных ингибиторов PTP-1B,
которые могут быть подходящими кандидатами для лечения диабета 2 типа и ожирения. Подробнее об этом можно узнать из оригинальной статьи, перейдя по
ссылке.
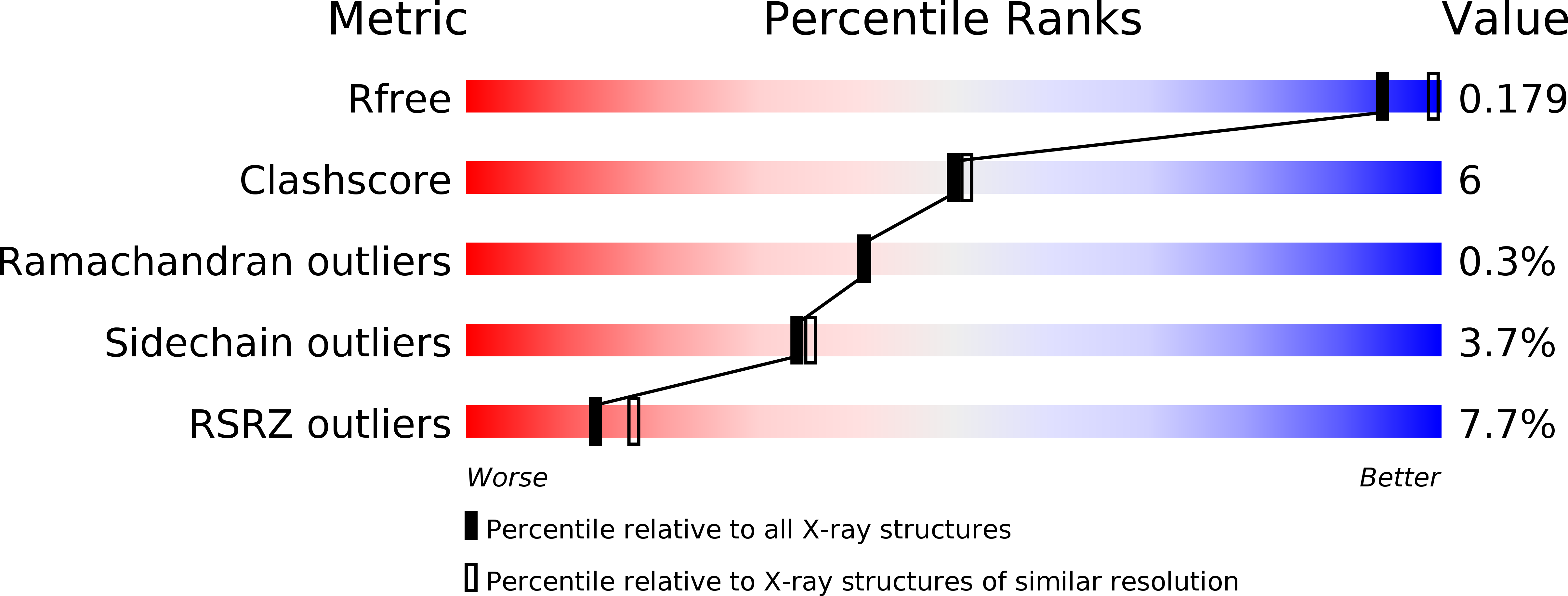
Рис.1 "slider" graphic с метрикой, сравнивающей ингибиторный комплекс PTPN1-inh с белками, которые ранее чем наш были депонированы в PDB.
По каждому показателю качества (всего 5) приводится число (в процентах) записей PDB, структуры которых не лучше структуры PTPN1-inh.
Если мы брали и сравнивали наш белок со структурами всевозможного разрешения, то полученное число наносили на слайдер в виде черной насечки, если близкого к разрешению у
PTPN1-inh - то в виде белой. Чем левее наносится засечка, тем соответственно меньше структур, которые нелучше по данному критерию чем наш белок.
По графику с метрикой понимаем, что показатель Rfree - один из показателей соответствия модели с тем, что пришло из эксперимента, хороший. Среди белков с похожим разрешением,
PTPN1-inh чуть ли не самый лучший, что замечательно, особенно если учитывать с каким высоким разрешением он был выделен. А по остальным показателям наш белок находится либо
в середине всего набора структур (согласно Clashscore, Ramachandran outliers, sidechain outliers), либо в хвосте (по RSRZ outliers). Последний показтель действительно настораживает, стоит ли авторам
изучать структурные особенности ингибиторного комплекса PTPN1 на белке 2ZMM, если почти его каждый 13ый остаток модели по своей электронной плотности больше чем на 20% отличается от экспериментальной?
Рис.2 Участок структуры PTPN1-inh, для которого нет подтверждающей его ЭП. Покрашен белок по B-фактору: неподвижные участки синие, а самые подвижные участки показаны красным и как
раз для них на нашлась ЭП, что закономерно и не вызывает вопросов, а наоборот наводит на мысль, почему белок 2ZMM по RSRZ показателю причислен к аутсайдерам. Не исключено, что таких
остатков с полярными группировками достаточное количество располагается на поверхности белка и следовательно все они колеблются в растворе, отчего вокруг них плохо описывается ЭП,
и они становятся маргиналами по RSRZ и тянут вниз по этому показателю белок PTPN1-inh.
2 задание
Пришло время узнать, какие именно остатки PTPN1-inh являются маргиналами и с чем это связано. В этом нам помог MolProbity, который сделал анализ геометрии структуры для лишенного водородов и
нашего белка и затем вновь их приобретшего, и выдал после этого соответствующую сводную таблицу по остаткам, где маргиналы были выделены красным и некотрые из них занесены в таблицу ниже.
Type of outliers
Outliers
Ramachandran
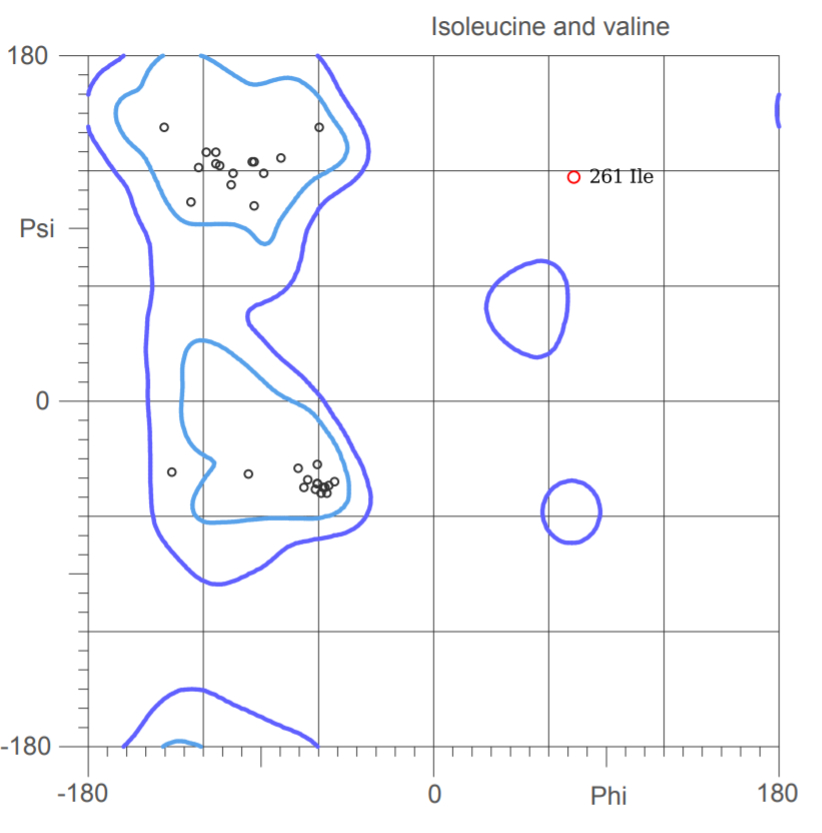
261 Ile
Rotamer
272,233,234,172 Leu , 11 Asp
Clashscore
3 Met, 246 Ile
Flip outliers
111 Asn, 157 Gln
Рис.3 Карта Рамачандрана для остатка Ile 261; чья пара углов Phi и Psi резко отличаются от предпочтительных значений.
Что касается маргиналов по торсионам боковых цепей, то здесь нет значимого влияния с их стороны на взаимодействие ингбитора с PTPN1, потому что эти маргиналы не связаны с функционально значимыми остатками и сами таковыми не являются.
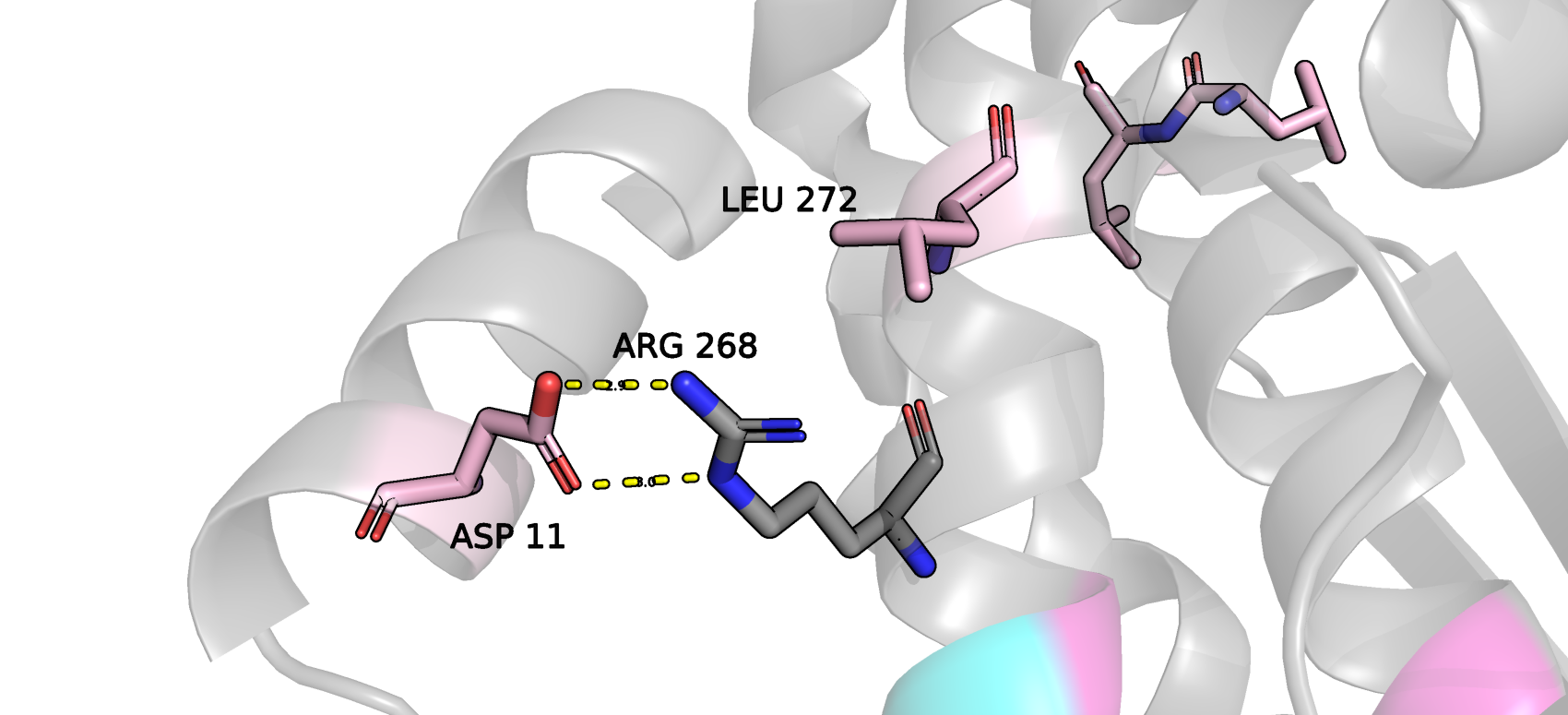
Стоит признаться, я не вижу никаких далеко идущих последствий их маргинальности, они вполне хорошо взаимодействуют с окружающими их остатками. На картинке ниже показан Asp11,
который соединен водородными связями с Arg268 и не требует валидации своего положения.
Рис.4 Положение в пространстве маргинала по торсионам Asp 11. Желтыми пунктирами выделены водородные связи Asp11 с Arg268.
Остаток Met3 локализован на конце альфа-спирали. MolProbity считает, что его Ван-дер-Ваальсов радиус перекрывается с таковым у Ile 246, который
находится в участке, где альфа-спираль теряет свою структуру, престает быть жесткой и приобретает подвижность, которая вероятно сделала эти 2 остатка
маргинальными. Но на взаимодействие ингибитора с PTPN1 эти остатки вряд ли влияют, они оба далеко расположены от активного центра.
Рис.5 Положение в пространстве маргиналов типа clash ILE246 и MET3.
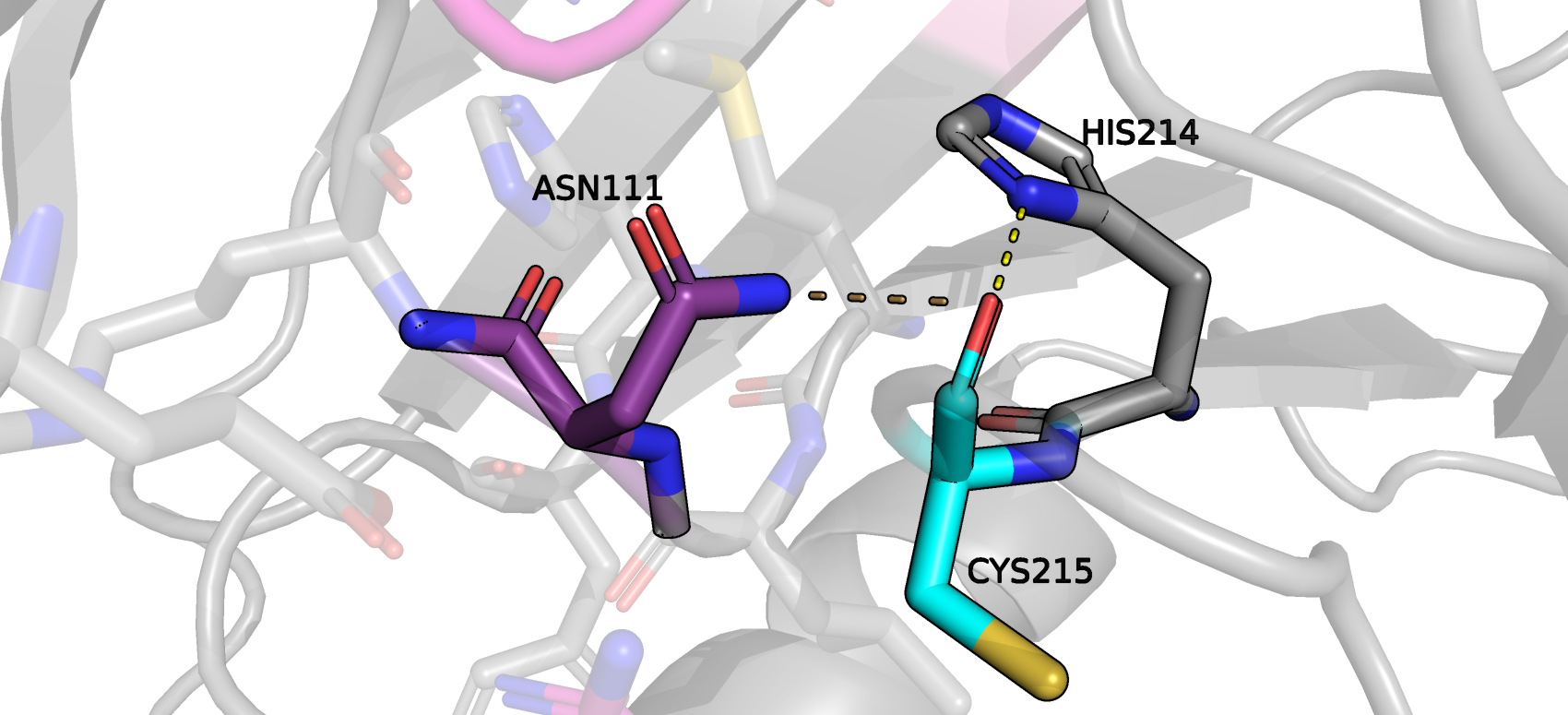
Остаток ASN111 был причислен к маргиналам, требующим разворота бокового радикала. И действительно, на рис.6 видно, что остаток будет комфортно находиться в своем окружении, если повернется и
образует водородную связь с карбонильным кислородом CYS215. Мне кажется, что ASN111, будучи маргиналом не будет нарушать связь между лигандом и CYS215, входящим в состав каталитического сайта. В итоге ни один
из маргиналов не лежит в функционально важном участке и не играет важной роли в образовании ингибиторного комплекса, что наряду с хорошим значением Rfree позволяет использовать данную структуру для изучения игибиторного
комплекса.
Рис.6 Положение в пространстве маргинала ASN111, требующего разворота. Он действительно этого требует, иначе не образуется водородная связь между азотом амидной группы
и кислородом остова CYS215. Ярко-желтый пунктир обозначает действительную водородную связь между CYS215 и HIS214, а коричневый - недействительную из-за неправильного положения боковой группы ASN111.
Рис.7 Структура PTP1B с ингибитором- производным 5-bromo-3-hydroxythiophene. Показаны
пунктиром водородные связи между карбонильной группой мочевинной части лиганда и водой, и между ней с ARG24. Эти связи а также циклогаксановая часть ингибитора, помещенная в карман PTP1B,
делают образующися комплекс устойчивым и таким образом оказывают ингибирующее действие на PTP1B. Известно, что рядом с этой связью отсутствуют какие-либо маргинальные остатки, что делает данную структуру возможной
для изучения данного комплекса.
4 задание
PDB-Redo использовался для улучшения качества нашей модели. Изменились в лучшую сторону все метрики по валидации:
Rfree, Ramachandran plot appearance, параметр, оценивающий правильность упаковки модели и др. Была удалена 31 молекула растворителя, для 69 остатков
PDB Redo значительно улучшил качество вписанности в электронную плотность (среди маргиналов таковыми остатками являются
почти все ротамерные маргиналы и 261Ile - Рамачандровский маргинал), и не было такого остатка, который стал бы хуже вписываться.
Переделанная модель не сильно отличается от исходной модели, если смотреть на их мультиковые отображения.
Рис.8 Представление в виде cartoon исходной (серый) и полученной вновь (голубой) моделей ингибиторного комплекса PTP1Binh. Видно, положения остова у двух можделей не сильно отличается.
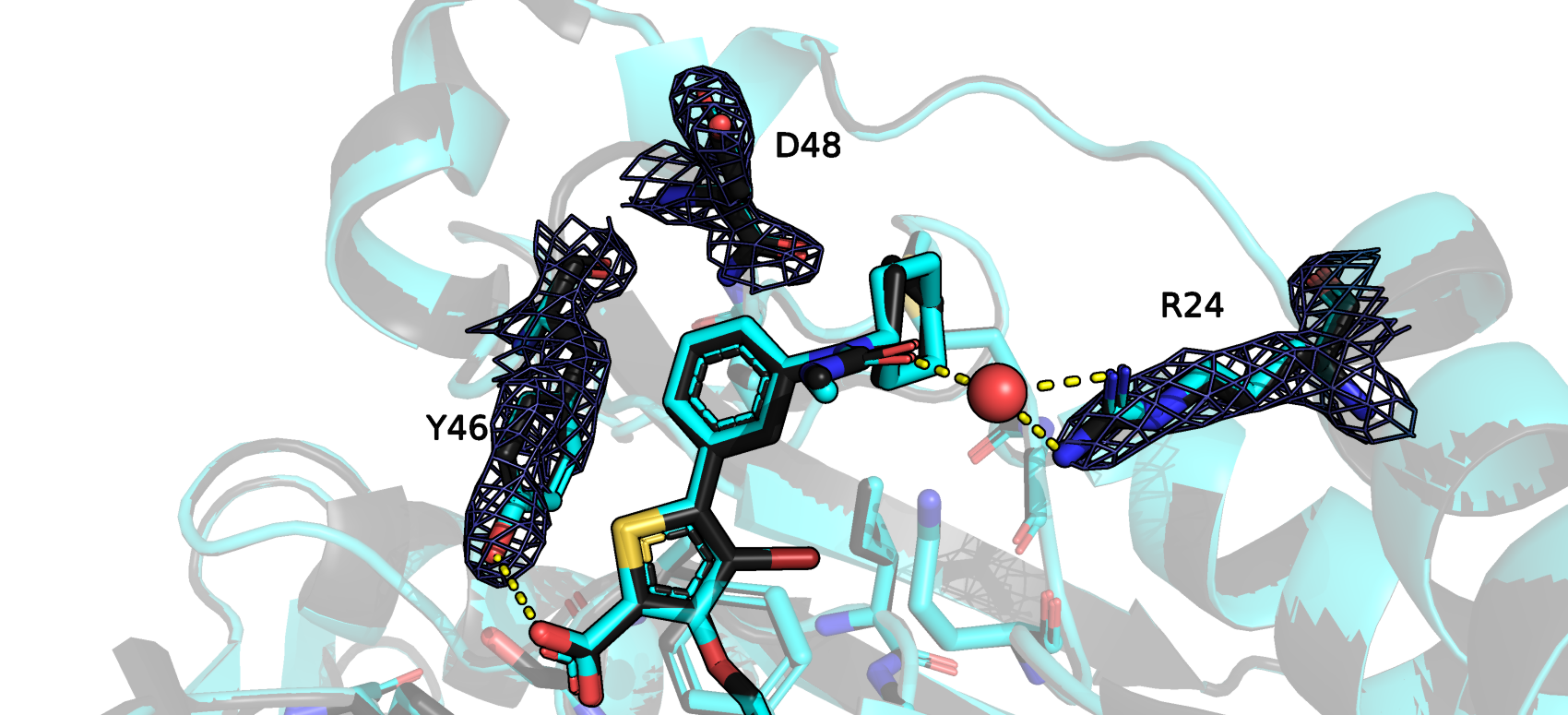
Было обращено внимание на остатки, участвующие в связывании лиганда с белком и чья вписанность в эп была резко улучшена Redo. Опосредованная водой
связь между мочевинной частью лиганда и Arg24 осталась неизменной, и качество вписанности также приемлемо как в исходной, так и в новой моделях. То же
можно сказать и про другие остатки, находящиеся в кармане связывания, поэтому ничто явно нам не мешает взять для исследования как исходную так и новую модель.
Рис.9 Представление в виде sticks некоторых остатков сайта связывания с лигандом у исходной (серый) и полученной вновь (голубой) моделей ингибиторного комплекса PTP1Binh.
Разница в том, как вписаны изображенные остатки новой и исходной моделей в электронную плотность, не велика. Водородная связь между остаком ARG24 и водой, между водой и лигандом, не изменялась Redo.