Постараемся в ручном режиме определить разметку неоднозначно проаннотированных остатков ингибиторного комплекса тирозинфосфотазы PTPN1-inh, для которого мы проводили анализ
на пригодность его модели (PDBid 2zmm) для работы по изучению его структурных и функциональных свойств.
1 задание.
Для разметки белка использовались программы DSSP и STRIDE. Обе программы приходили к консенсусу при разметке остатков, принадлежащих к элементам типа H(helix), E(betaStrand),
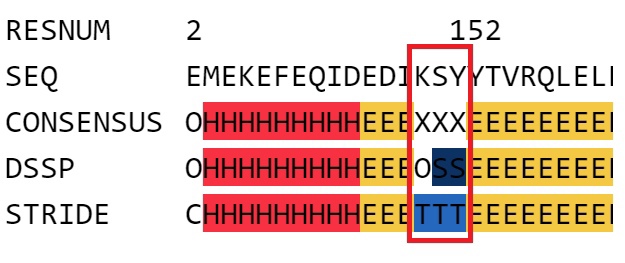
но расходились во мнениях при разметке остатков, расположенных в петлях. В качестве примера приведены следущие друг за другом остатки K150,S151,Y152.
Рис.1 Фрагмент белка PTPN1-inh, который был по-разному размечен программами DSSP и STRIDE. DSSP не разметила K150 (символ O), последующие за ним остатки приписала излому (S).
STRIDE решил, что эти остатки приходятся на поворот, который, будь он побольше, образовал бы спираль.
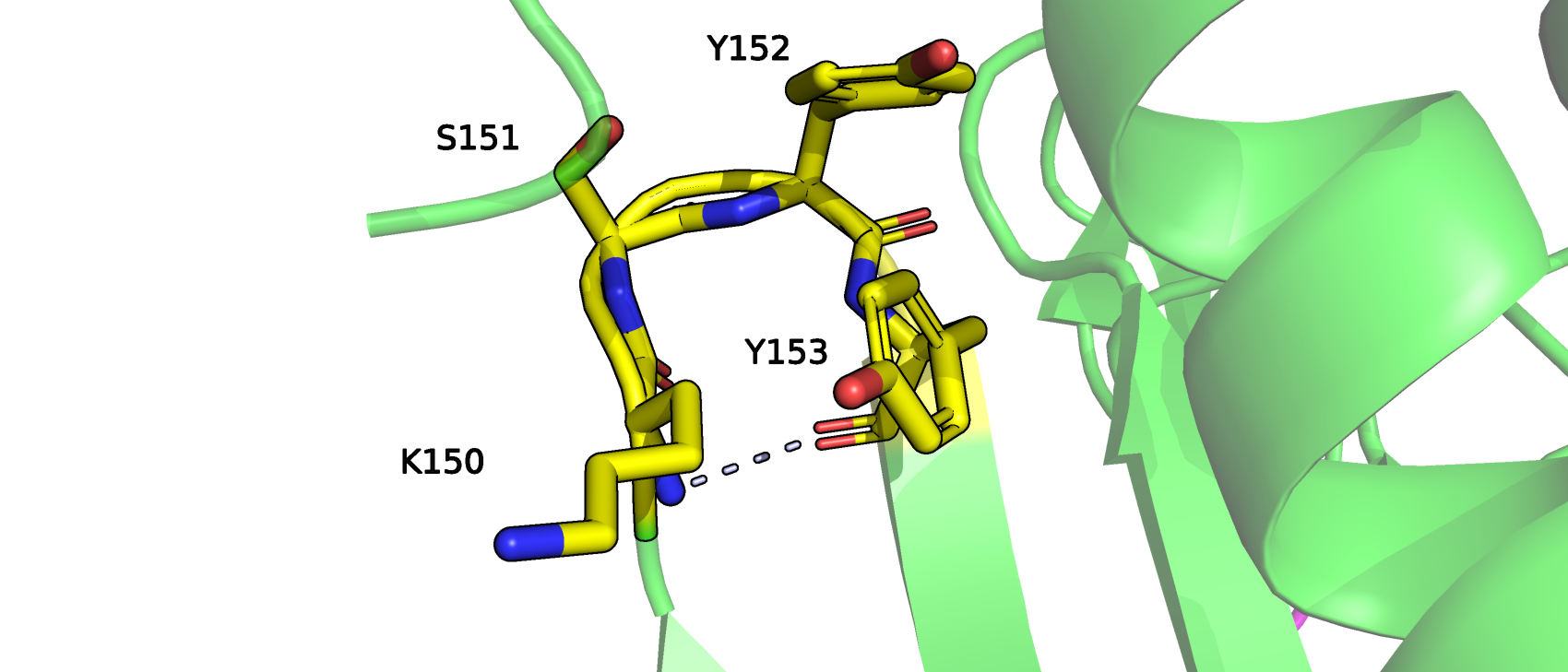
Увидев эти остатки в паймоле, мы решили согласиться насчет лизина с STRIDE. Его остовынй азот образует водородную связь с остовным кислородом остатка Y153 (рис.2), следующего за ним через 2 другие аминокислоты (S151, Y152).
Последних мы относим к излому, то есть соглашаемся с версией DSSP, потому что в этом месте меняется направление остова на противоположное.
Рис.2 Визуализация в Pymol фрагмента белка PTPN1-inh, который был по-разному размечен программами DSSP и STRIDE. Голубым показана водородная связь между K150 и Y153 - признак 3-поворота.
Мне не удалось найти связи между особенностями алгоритмов разметки с их вердиктом для рассмотренных остатков. Но я думаю, что DSSP могла согласиться со STRIDE насчет 3-поворота для лизина K150, увидев
водородную связь между ним и Y153 (чего она не сделала, видимо потому что рассчитывает E водородной связи проще чем STRIDE). И поэтому не буду исключать, что разница в подсчете энергии водородной связи привела
к разной разметке лизина K150.
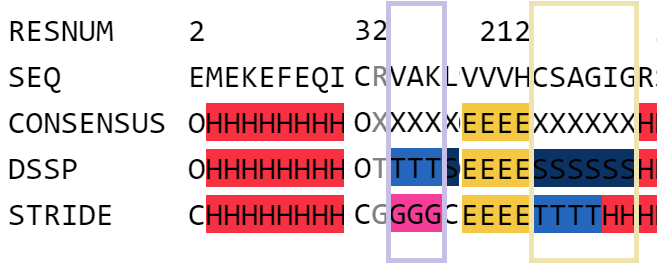
Рассмотрим еще 2 по-разному размеченных фрагмента. На рис.3 первый фрагмент (с 33-го по 36ой остатки, обведен в сиреневую рамку)
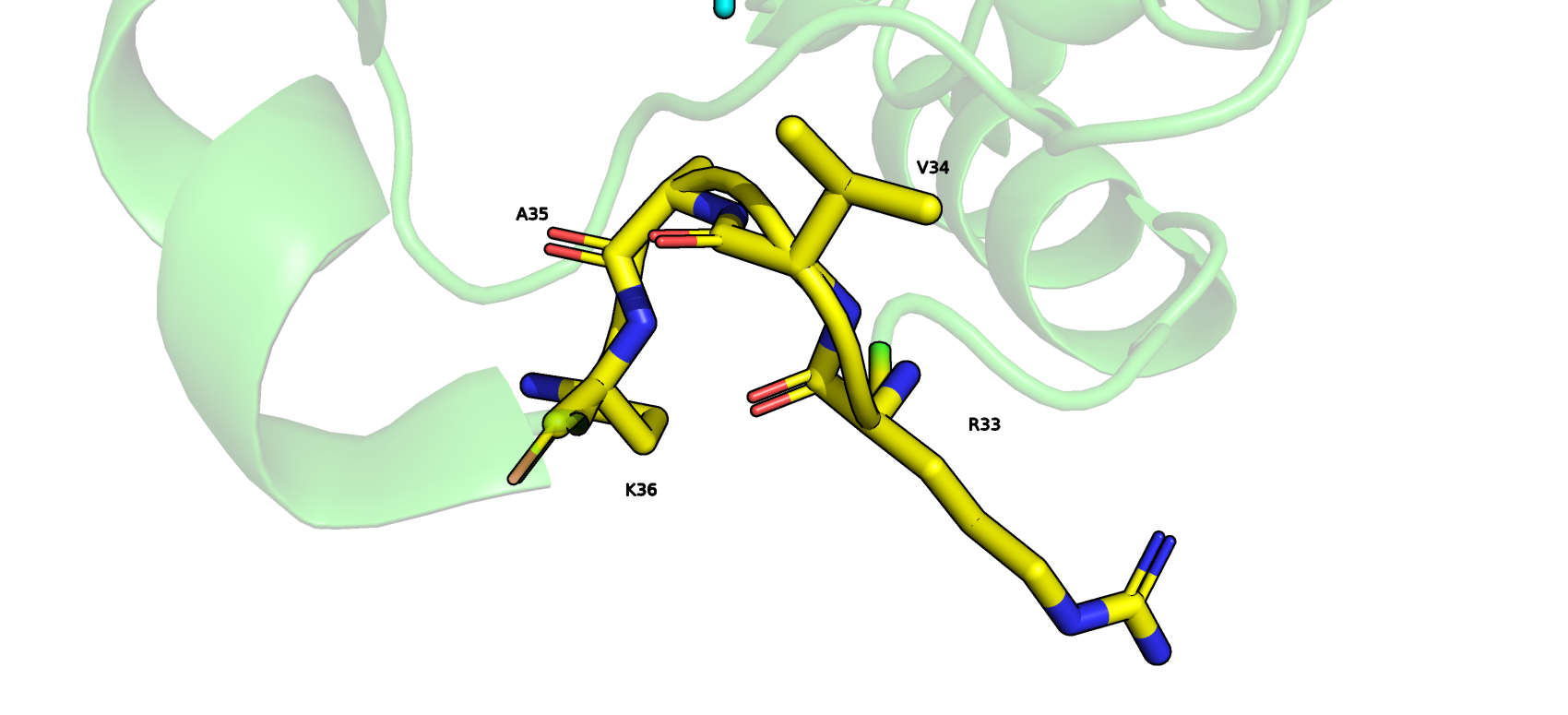
по версии DSSP представлен 3-поворотом, а по версии STRIDE- это полноценная 3/10 спираль. Визуализация в pyMol (рис.4) не согласуется с обеими версиями, там
отсутствуют водородные связи между остовными атомами азота iго и кислорода i+2го остатков, поэтому данные фрагменты не являются элементами 3-поворота и тем более
3/10 спирали. Я бы отнес рассматриваемые остатки к изгибу. Здесь меняется направление остова на угол, больший чем 70 градусов.
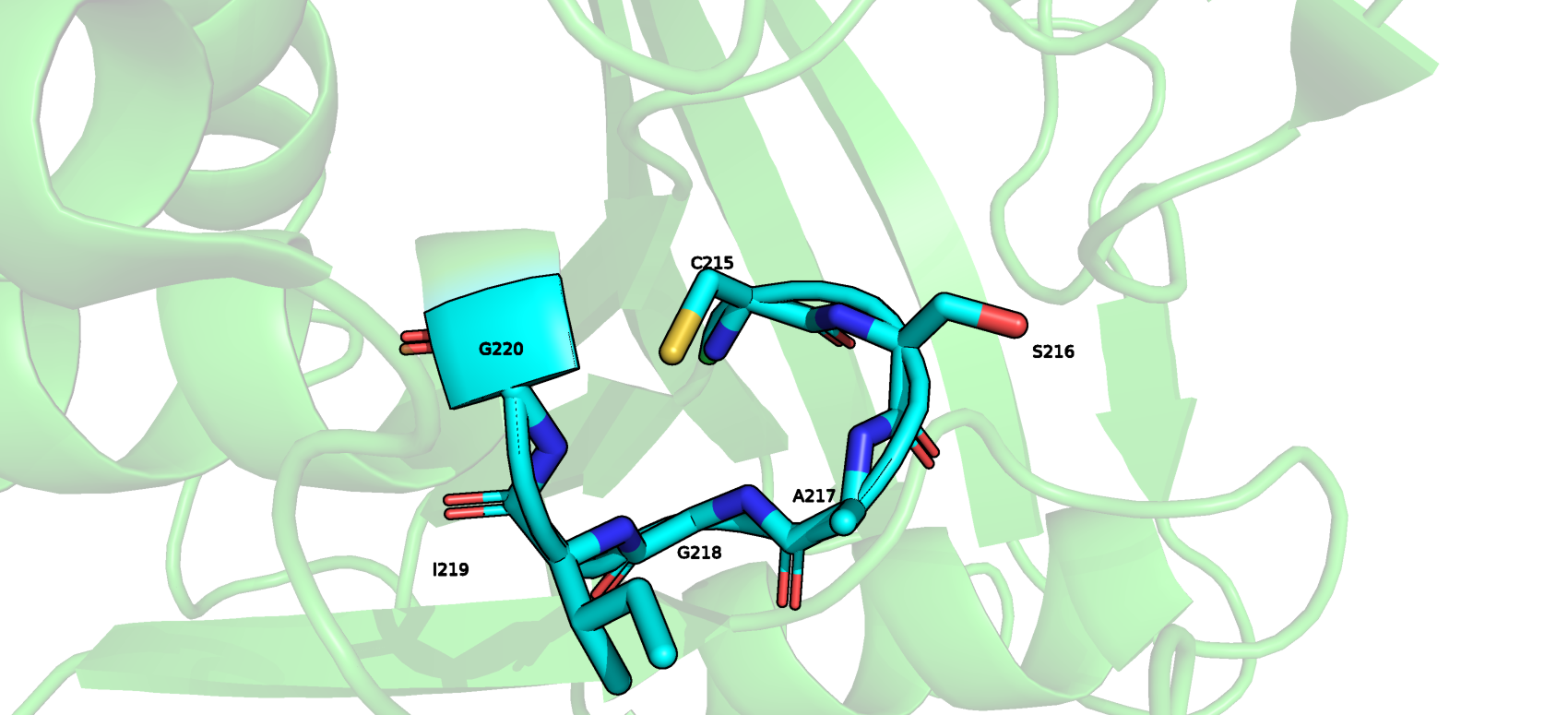
На том же рис.3 второй фрагмент (с 215го по 220ый, обведен в персиковую рамку) по версии DSSP представлен изгибом (S), а по версии STRIDE- он представлен сначала
3-поворотом, который на конце сменяется остатками альфа-спирали. Этот фрагмент, точнее его первые 5 остатков, я бы тоже, как и в предыдущем случае, отнес к изгибу.
На рис.5 у этого участка отсутствуют описанные выше водородные связи, зато направление его остова круто меняется, как это происходит в изгибе. А насчет последнего
остатка я согласен со STRIDE, я бы отнес G220 к альфас-спирали, он и в PуMol сидит на спирали.
Рис.3 Фрагменты белка (с 33-го по 36ой, обведен в сиреневую рамку) и (с 215го по 220ый, обведен в персиковую рамку) PTPN1-inh, которые были по-разному размечены программами DSSP и STRIDE.
Рис.4 Визуализация в Pymol фрагмента (с 33-го по 36ой остатки) белка PTPN1-inh, который был по-разному размечен программами DSSP и STRIDE.
Здесь отсутствуют водородные связи между остовными атомами азота iго и кислорода i+2го остатков, поэтому данные фрагменты не являются элементами 3-поворота и тем более
3/10 спирали.
Рис.5 Визуализация в Pymol фрагмента (с 215-го по 220ый остатки) белка PTPN1-inh, который был по-разному размечен программами DSSP и STRIDE.
2 задание.
Решили узнать, имея в руках несколько файлов pdb, какие остатки склонны преимущественно находиться в бета-тяжах(E), спиралях(H) и других структурах (C).
В итоге получили таблицу(смотрите ниже) с показателями склонности каждого остатка образовывать тот или иной тип вторичной структуры. Выяснилось, что Валин чаще образует бета-тяжи, глутамат преимущественно расположен в альфа-спиралях.
Аспарагин, аспартат, глицин и пролин склонны находиться в петлях. Могу прокомментировать посчитанные показатели для пролина: остовный азот остатка лишен водорода, с помощью которого он образует водородную связь
с кислородом другого остатка для поддержания в пространстве спирали. Отсюда его редкое появление в данного типа вторичной структуре и соответсвенно более частое пребывание в петлях, где атомам остова водородных связей между собой
образовывать необязательно.
E H C
A 0.868 1.314 0.819
R 0.956 1.103 0.941
N 0.449 0.725 1.452
D 0.514 0.641 1.488
C 1.289 1.006 0.868
Q 0.91 1.143 0.931
G 0.699 0.449 1.553
E 0.646 1.318 0.913
H 1.058 1.103 0.896
I 1.686 1.012 0.689
L 1.012 1.404 0.687
K 0.838 1.173 0.939
M 0.989 1.39 0.707
F 1.109 0.953 0.988
P 0.513 0.521 1.58
S 0.894 0.741 1.245
T 1.221 0.901 0.978
W 1.251 0.938 0.936
Y 1.49 1.117 0.695
V 1.996 0.868 0.662