Предисловие: в дополнительном задании я использовал белок с id 2GVV, так как у назначенного мне белка отсутвует статья.
Третий практикум
Структура в целом
Данный белок состоит из одной макромолекулы, являющейся потенциальным тиолом. Есть 4 одинаковых цепи, входящих в единственную макромолекулу.
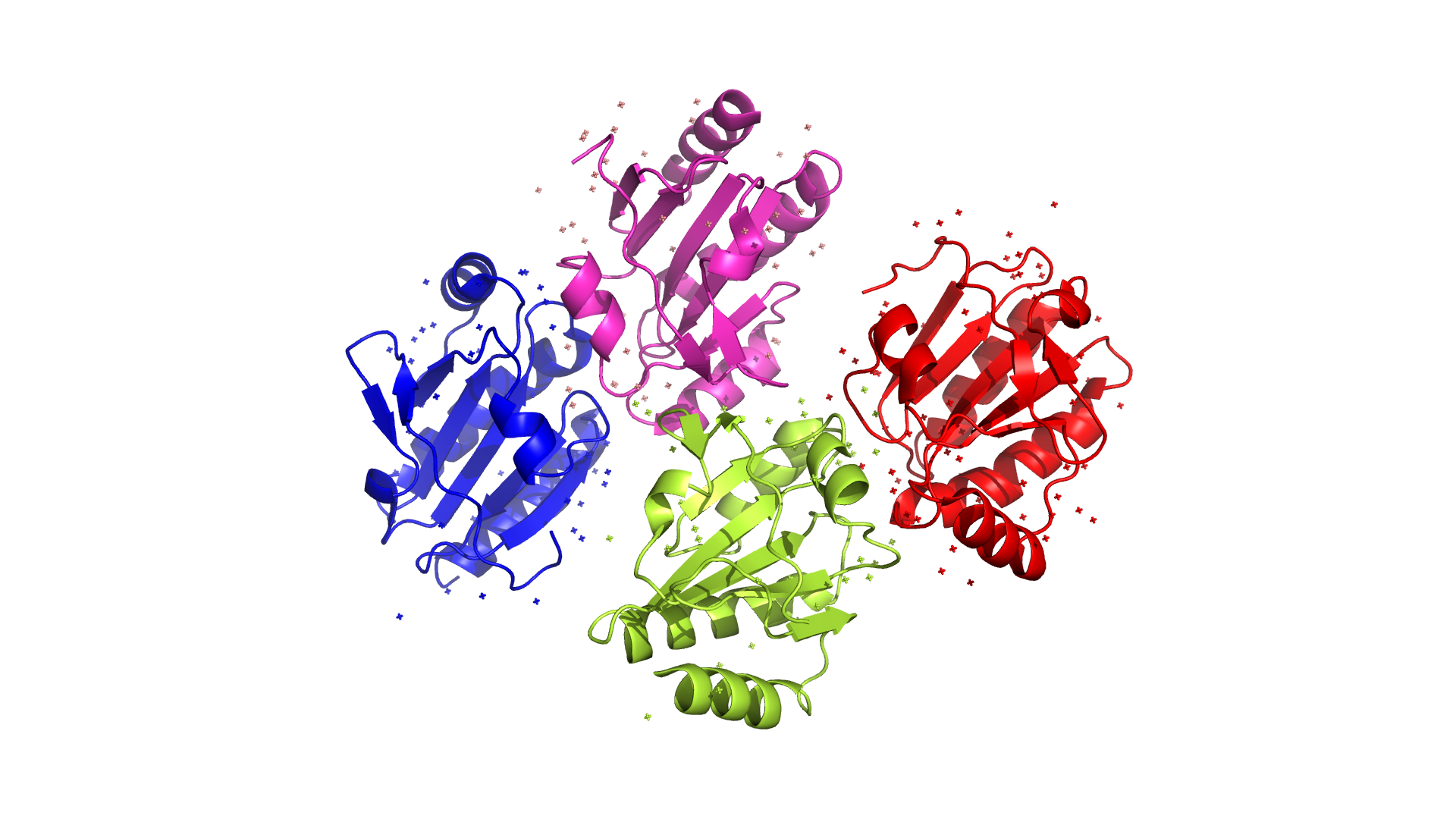
Отдельные цепи
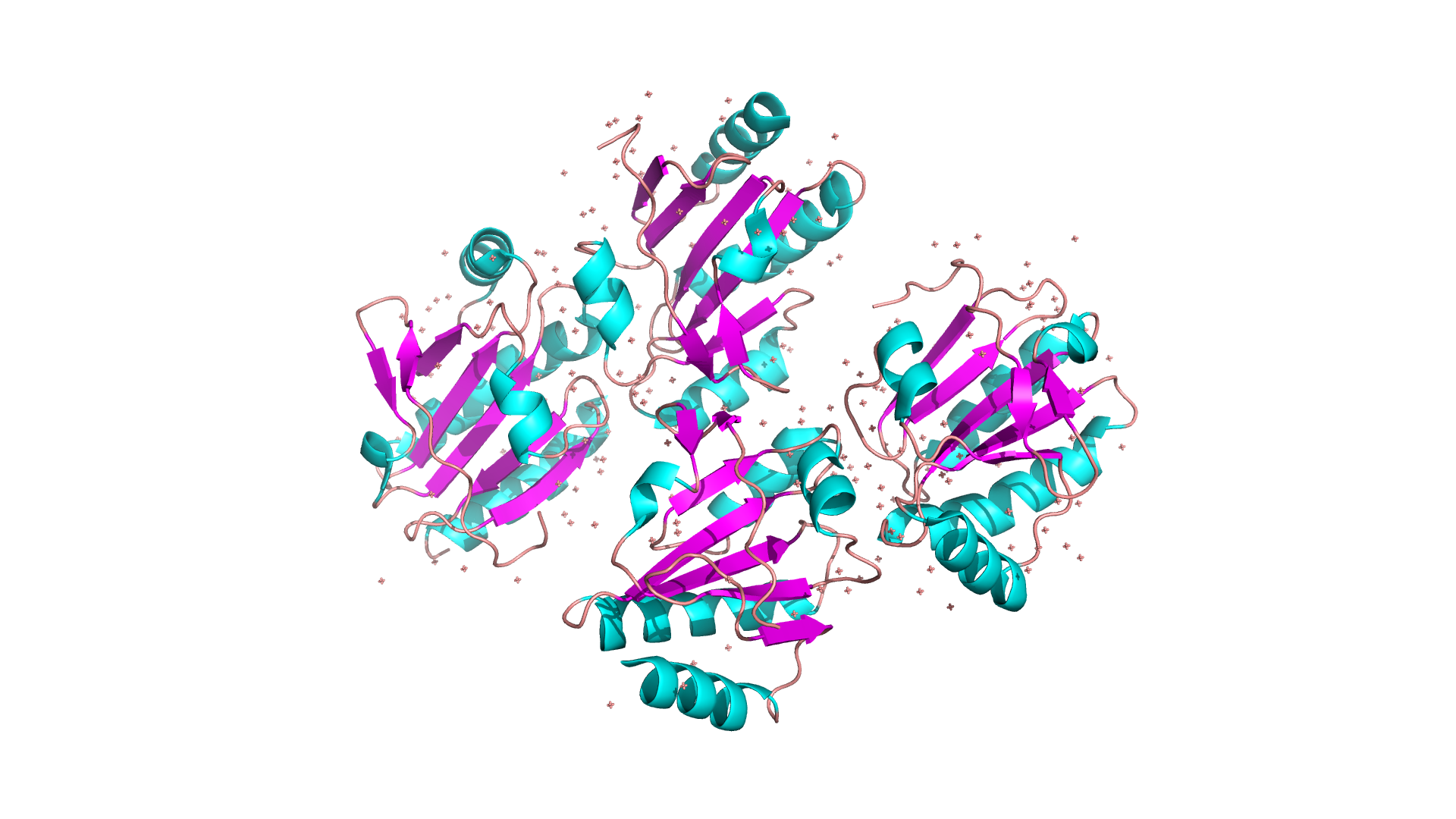
Единственная макромолекула белка принадлежит бактерии Chlorobaculum tepidum TLS (зеленая серная бактерия). Uniptot ID: Q8KDH8, мутаций нет. При этом есть 16 модифицированных аминокислот, все из которых являются метионином. Файл, содержащий строки с информацией: *ссылка*.
Малые молекулы
В белке, по заявлению страницы на pdb, присутвует всего один тип малых молекул - selenomethionine (MSE), присутвующих во всех цепях белка. Однако в файле формате pdb я нашел так же заявление о присутствии воды (HOH) в молекуле. Информация о малых молекулах в файле .pdb: *ссылка*
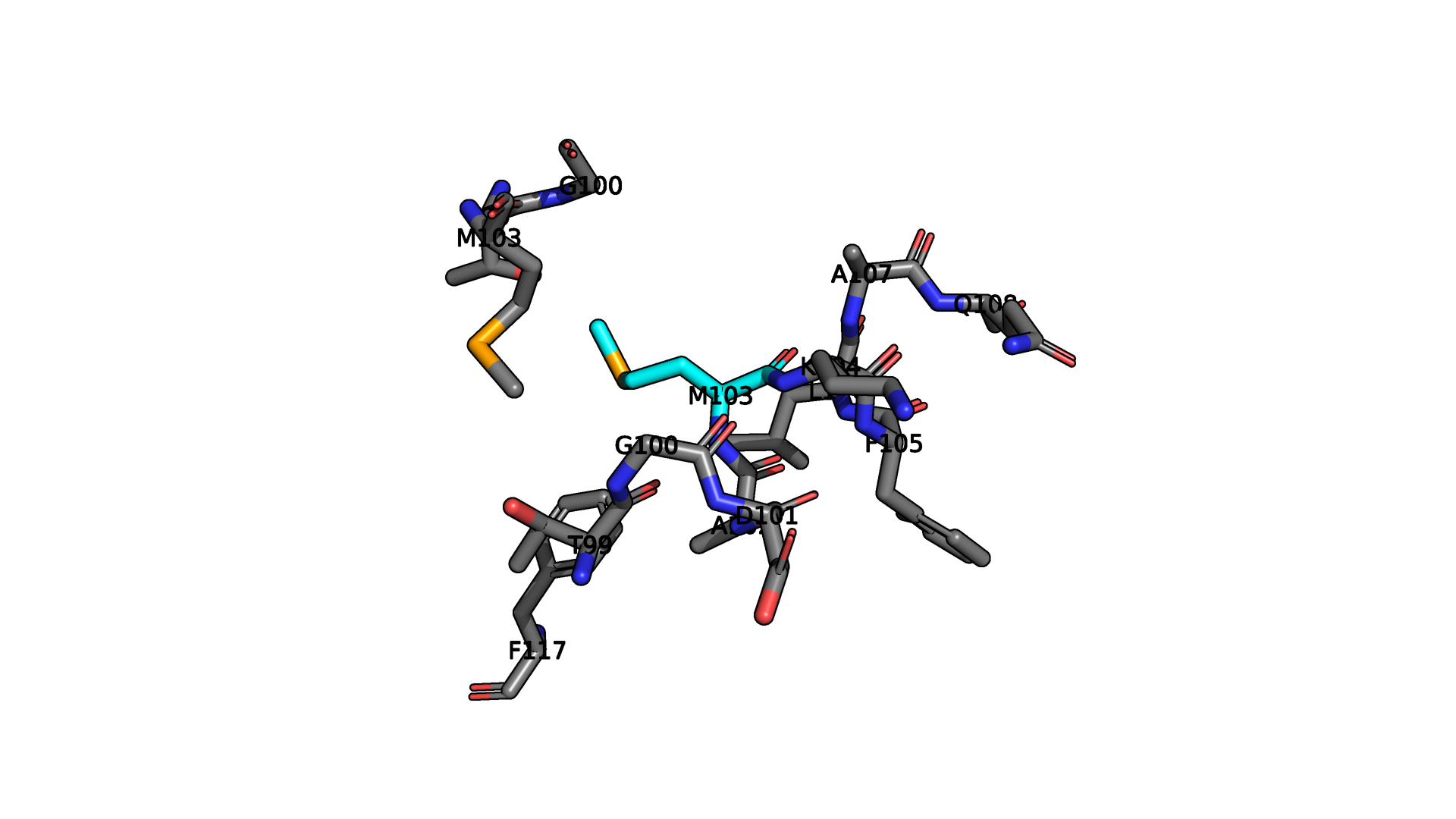
Взаимодействия между аминокислотными остатками
В данном белке пристувуют все типы взаимодействий, кроме дисульфидных мостиков. Примеры приведены ниже.
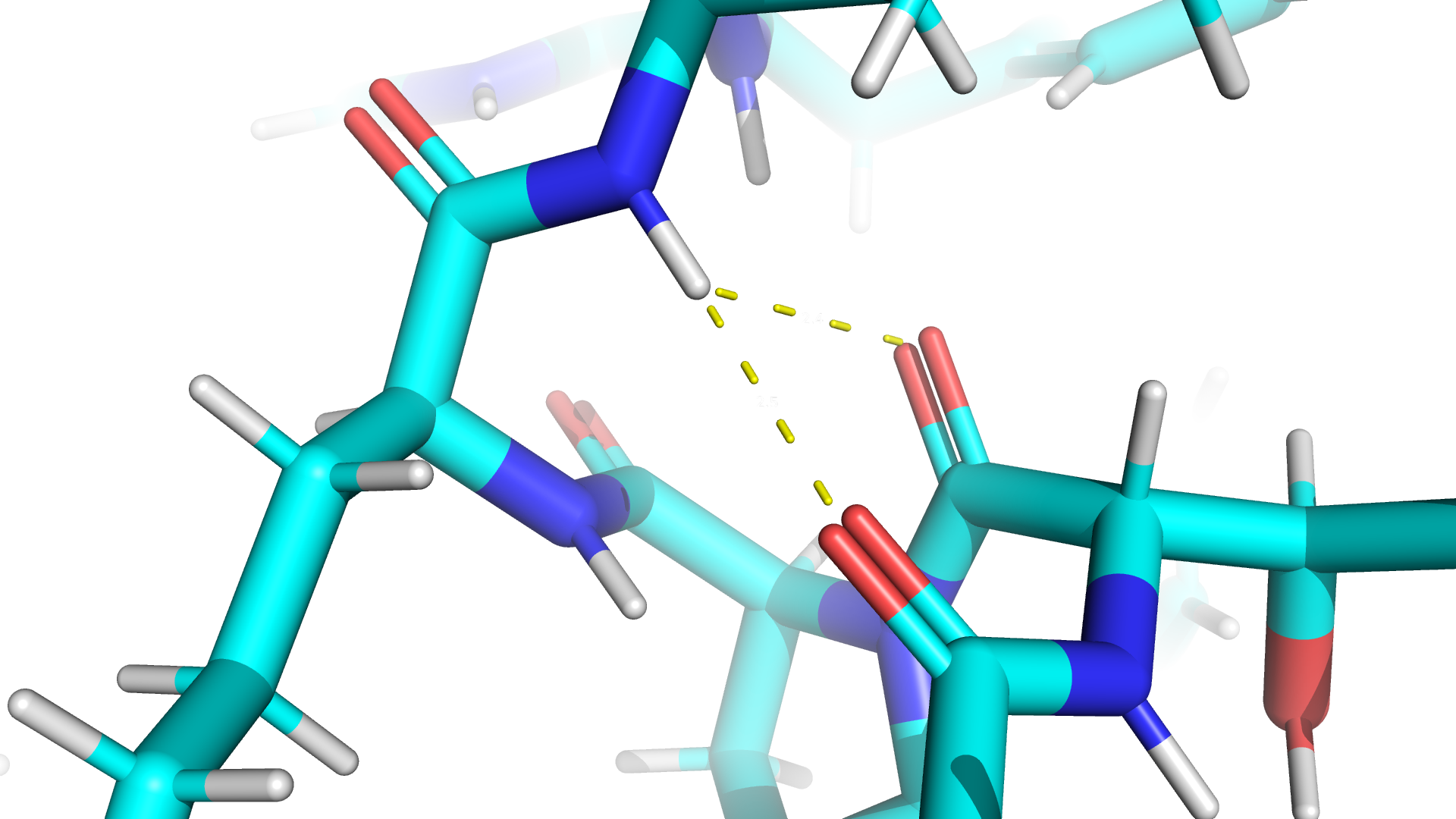
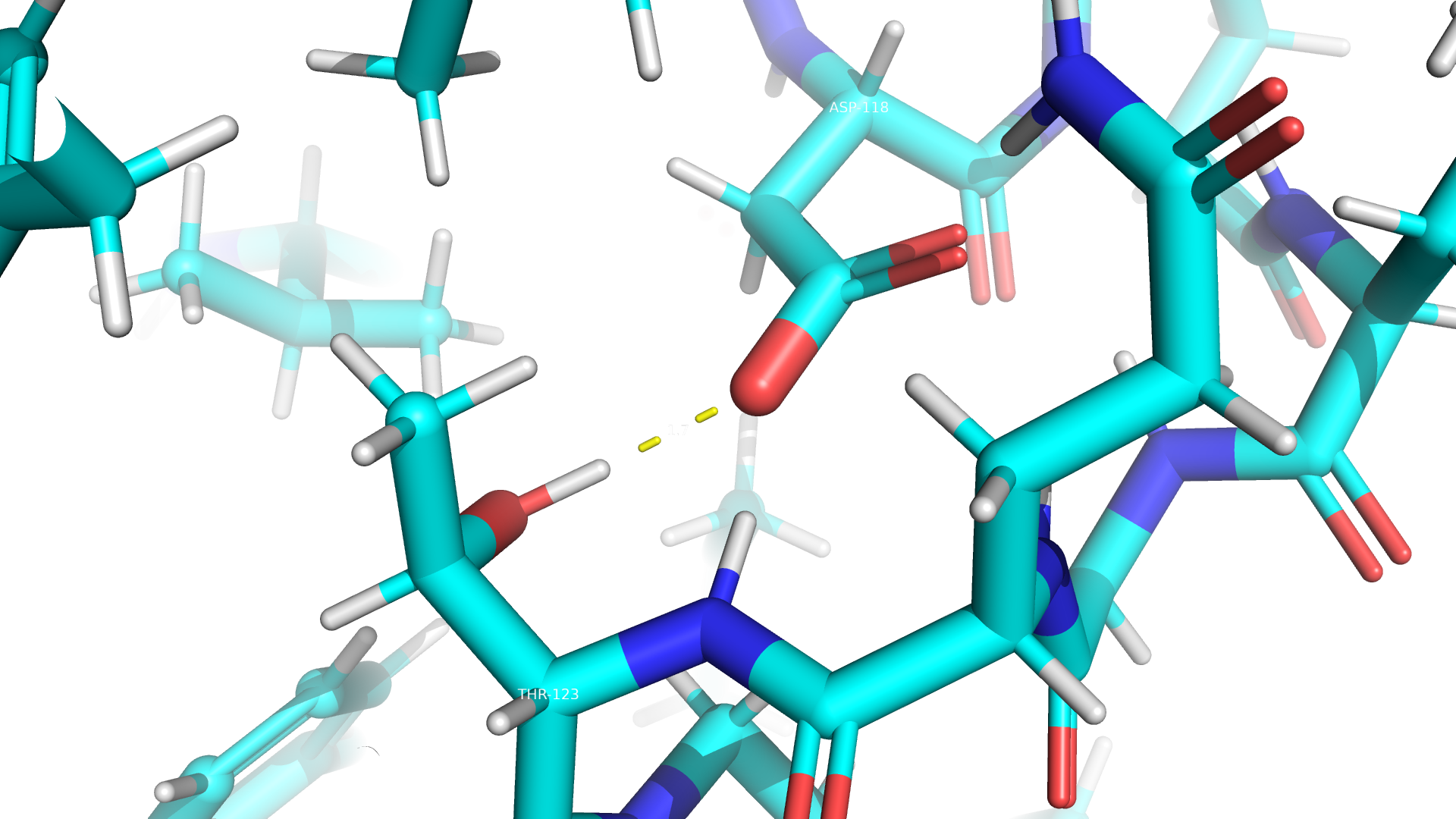
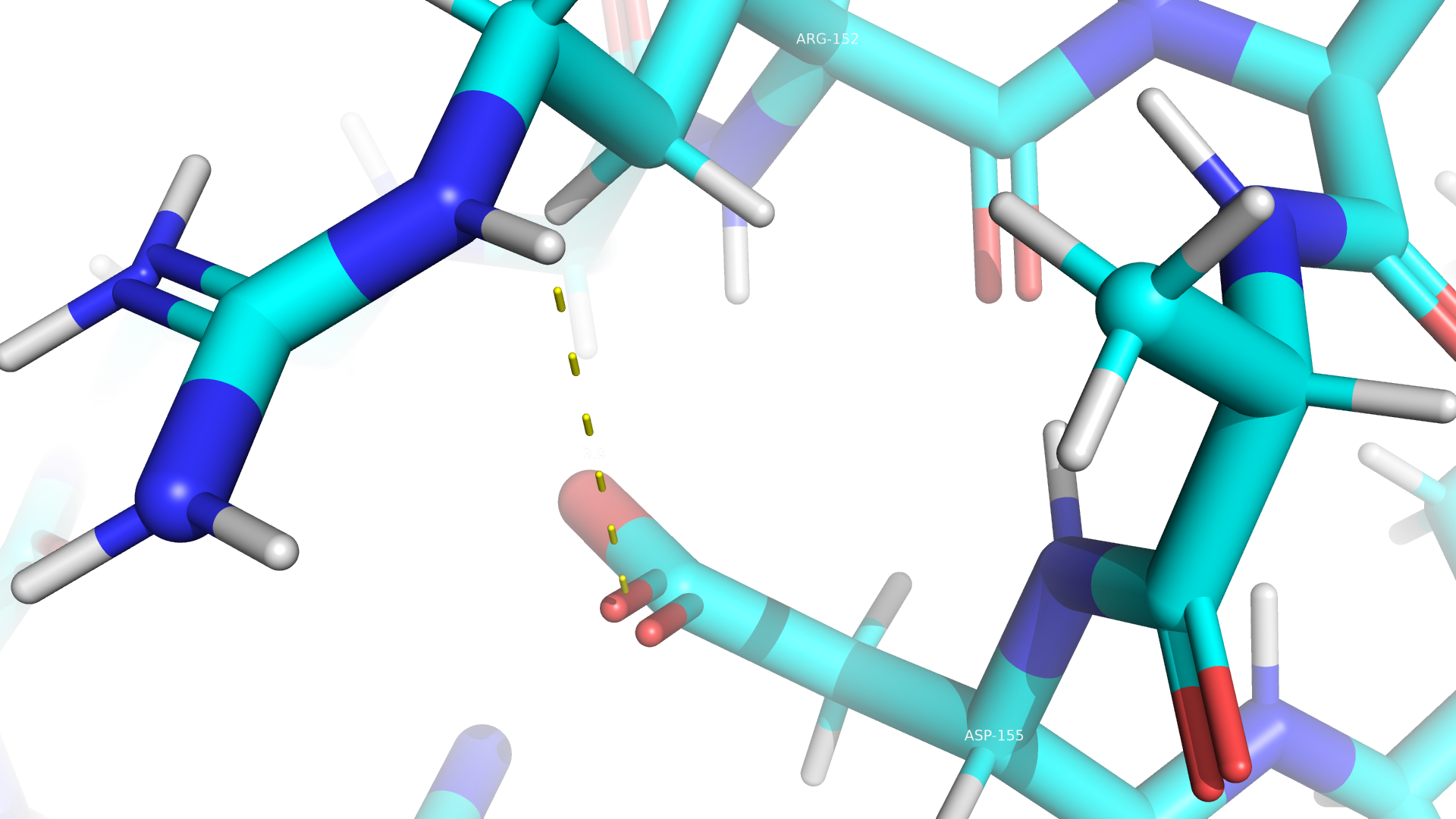
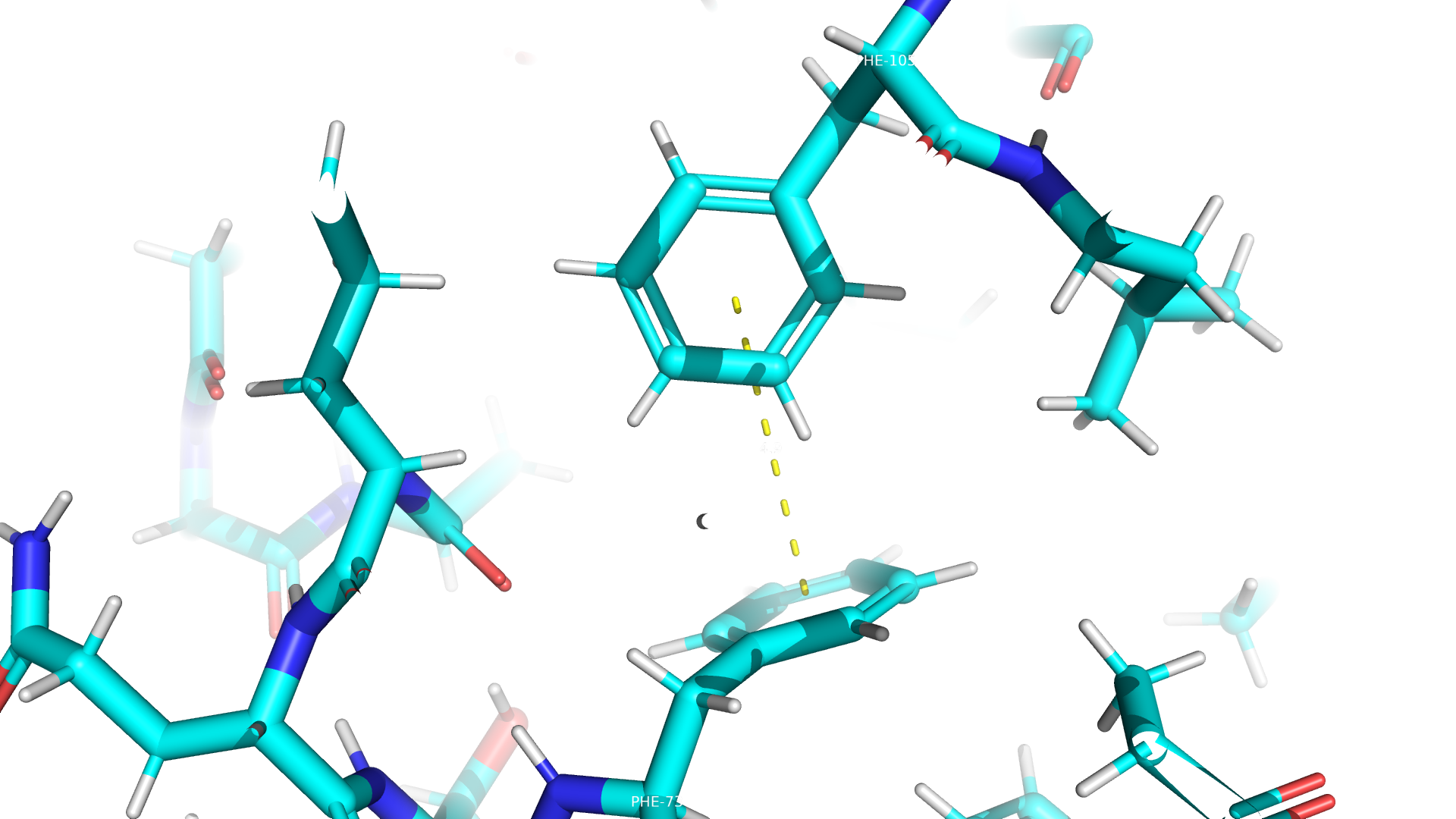
Дополнительное задание
К сожалению, статья о назначенной мне молекуле не выпущена, так что я буду использовать белок с pdb id 2GVV. В данном белке две малые молукулы: дициклопентилфосфорамидат (DI9) и ионы кальция (CA). DI9 служит для координации фосфорильного кислорода, и в качестве катализатора используется CA.