Учебный сайт Валяевой Анны
Комплексы ДНК-белок
Предсказание вторичной структуры заданной тРНК
Работа проводилась с нуклеотидной последовательностью тРНК 2DXI, скачанной из PDB банка.
С помощью программы find_pair были получены данные о комплементарных основаниях, по которым можно определить положение стеблей во вторичной структуре тРНК. Путем поиска инвертированных повторов с помощью программы einverted из пакета EMBOSS были найдены возможные комплементарные участки в последовательности исследуемой тРНК. В ходе анализа менялись значения gap penalty и minimum score threshold. В результате при gap penalty = 12 и minimum score threshold = 15 программа нашла однин инвертированный повтор, находящийся в акцепторном стебеле:
SEQUENCE: Score 15: 5/5 (100%) matches, 0 gaps
3 cccca 7
|||||
69 ggggt 65
При понижении gap penalty до 2 и minimum score threshold = 15 программа выдала более длинный участок, но содержащий много гэпов, поэтому его не учитываем при предсказании вторичной структуры тРНК.
Программа einverted проводит поиск только по каноническим парам, из-за этого он оказывается неэффективен.
Далее было проведено предсказание с помощью алгоритма Зукера, который реализуется программой mfold. Наиболее близкая к реальному структура получилась при P=10, она представлена на рисунке 1. При увеличении значения P количество возможных вторичных структур увеличивалось, но среди них появлялись структуры, не характерные для тРНК.
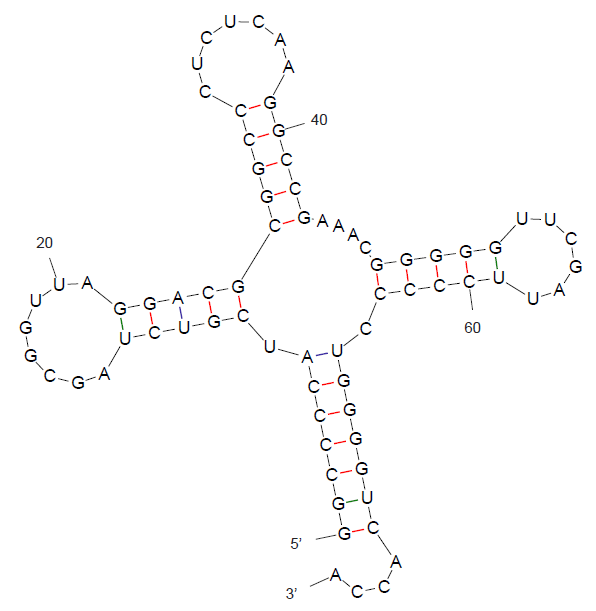
Рис. 1. Вторичная структура тРНК 2DXI, предсказанная с помощью алгоритма Зукера.
Таблица 1. Реальная и предсказанная вторичная структура тРНК из файла 2DXI.pdb.
| Участок структуры | Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
| Акцепторный стебель | 5'-501-507-3' 5'-566-572-3' Всего 7 пар | предсказано 5 пар из 7 реальных | предсказано 7 пар |
| D-стебель | 5'-510-513-3' 5'-522-525-3' Всего 4 пары | - | предсказано 5 пар |
| T-стебель | 5'-549-553-3' 5'-561-565-3' Всего 5 пар | - | предсказано 5 пар |
| Антикодоновый стебель | 5'-526-531-3' 5'-539-544-3' Всего 6 пар | - | предсказано 5 пар |
| Общее число канонических пар нуклеотидов | 20 | 5 | 22 |
Поиск ДНК-белковых контактов в заданной структуре
Для определения ДНК-белковых контактов в структуре комплекса из файла BY4.pdb был написан скрипт для Jmol. Будем считать полярными атомы кислорода и азота, а неполярными - атомы углерода, фосфора и серы. Назовем полярным контактом ситуацию, в которой расстояние между полярным атомом белка и полярным атомом ДНК меньше 3.5A. Аналогично, неполярным контактом будем считать пару неполярных атомов на расстоянии меньше 4.5A. Результаты представлены в таблице 2.
Таблица 2. Контакты разного типа в комплексе 1BY4.pdb.
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 5 | 17 | 22 |
| остатками фосфорной кислоты | 24 | 18 | 42 |
| остатками азотистых оснований со стороны большой бороздки | 4 | 11 | 15 |
| остатками азотистых оснований со стороны малой бороздки | 0 | 3 | 3 |
Как видно из таблицы, белок (Retinoid X receptor alpha) главным образом связывается с участком ДНК через сахарофосфатный остов и большую бороздку, тем самым влияя на транскрипцию.
Далее с помощью программы nucplot была получена популярная схема ДНК-белковых контактов. Она представлена на рисунках 2 и 3. Использованные команды:
remediator --old 1BY4.pdb > 1BY4_old.pdb nucplot 1BY4_old.pdb convert nucplot.ps nucplot.png
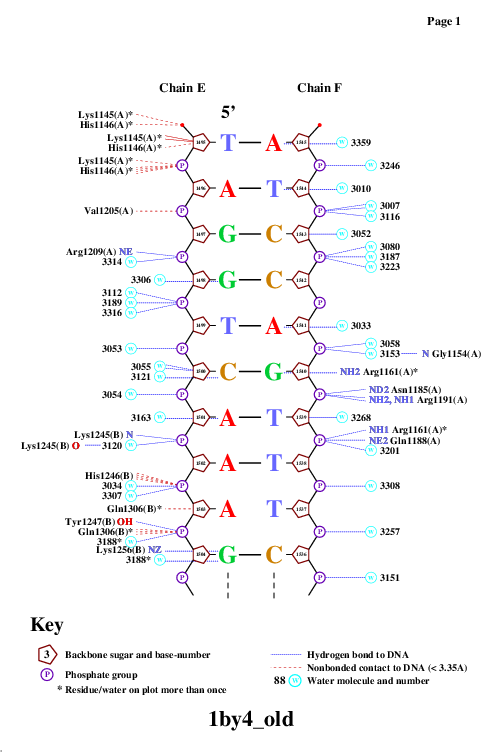
Рис. 2. Популярная схема ДНК-белковых контактов в структуре 1BY4. Изображение получено с помощью программы nucplot.
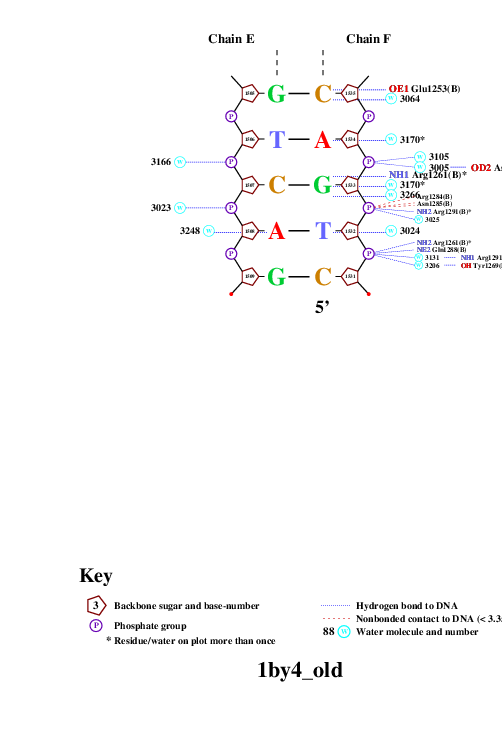
Рис. 3. Популярная схема ДНК-белковых контактов в структуре 1BY4. Изображение получено с помощью программы nucplot.
На схеме видно, что наиболее "загруженными" аминокислотными остатками (с наибольшим числом указанных на схеме контактов с ДНК) являются лизин 1145 и гистидин 1146. На рисунке 4 изображено взаимодействие Glu1253 и ДНК, которое, возможно, важно для распознавания последовательности.
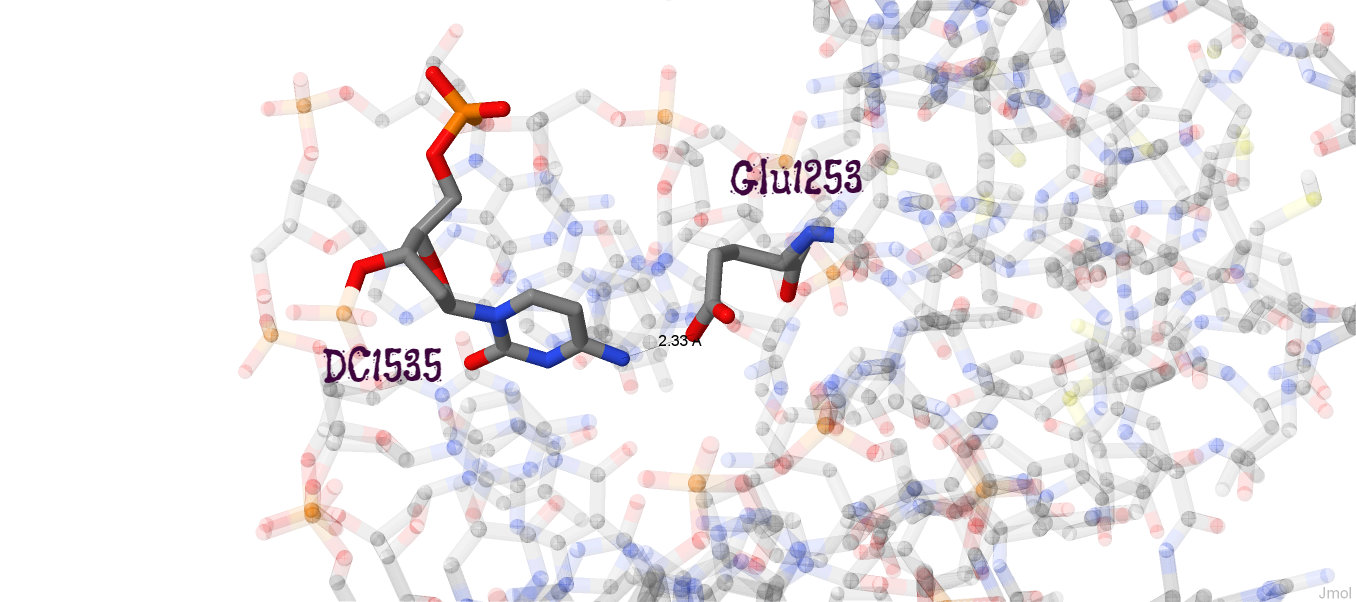
Рис. 4. Взаимодействие глутаминовой кислоты и цитозина ДНК. Изображение получено с помощью программы Jmol.
Дата последнего обновления: 01.12.14
©Валяева Анна