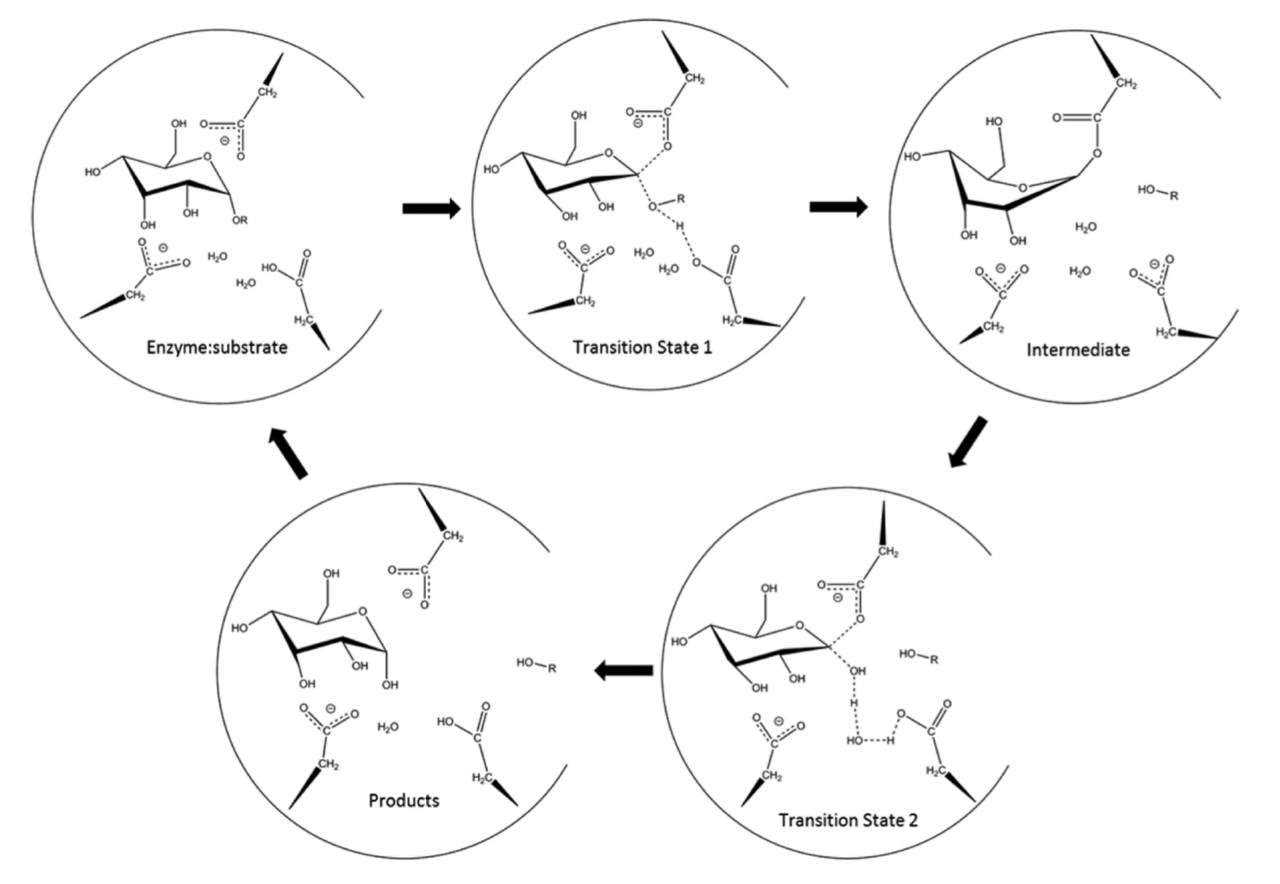
Данный фермент относится к семейству гидрозилгидролаз 13 [1]. Панкреатическая
альфа-амилаза осуществляет гидролиз альфа(1-4) гликозидных связей в полисахаридах. Активный центр данной
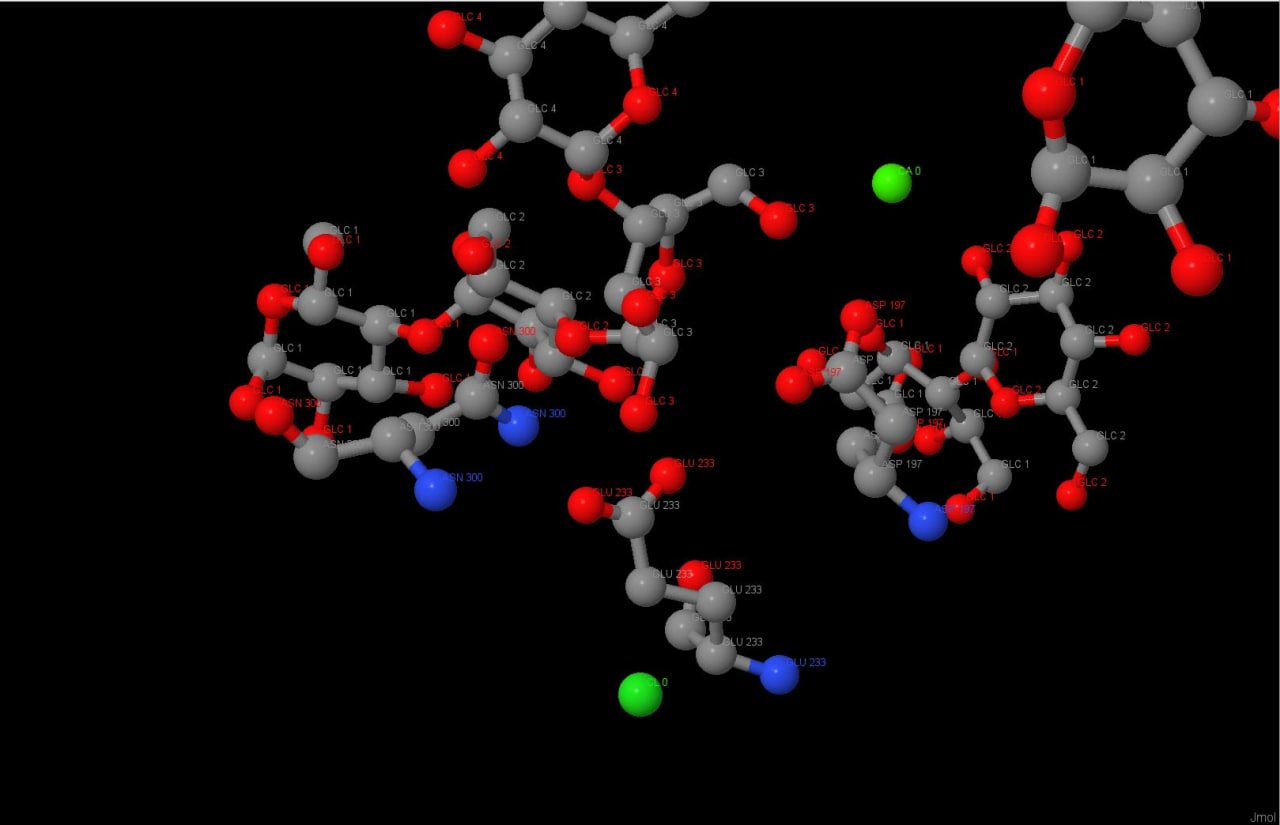
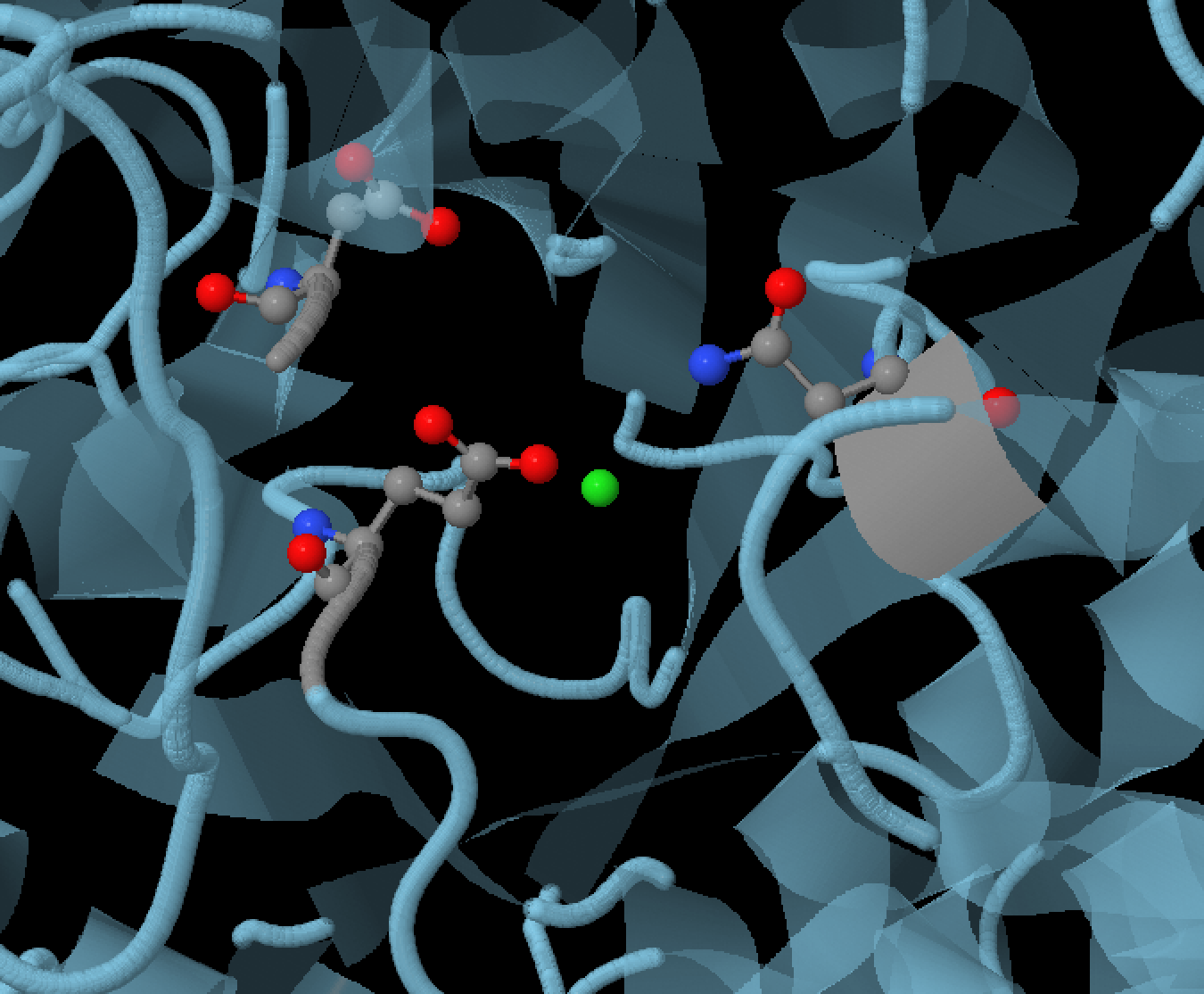
альфа-амилазы состоит из трех аминокислотных остатков: Asp300, Glu233 и Asp197 (Рисунок 2). Каталитическую реакцию
нашего фермента можно разделить на две стадии: гликозилирование и дегликозилирование. При гликозилировании
депротонированный Asp300 образует с двумя гидроксильными остатками одного из колец полисахарида водородные связи,
тем самым удерживает полисахараид в правильном положении для катализа. Водород карбоксильной группы Glu233 образует
с кислородном гликозидной связи водородную связь, также стабилизируя субстрат для дальнейшего катализа (стоит отметить,
что в отличии от других двух аминокислотных остатков, Glu233 не лишен протона и является в данной реакции его донором).
Кислород, который не соединен с водородом карбоксильной группы Asp197, соединяется с водородом гидроксильной группы
олисахарида (вероятно это также играет важную роль в поддержаниии правильного пространственного положения субстрата),
при этом второй кислород выходит на хорошую позицию для нуклеофильной атаки на аномерный углерод (соединен с кислородом гликозидной
связи). Важной частью данной каталитической реакции являются две молекулы воды. При стадии присоединения субстрата к
активному центру она находится рядом с Glu233. При нуклеофильной атаке Asp197 на аномерный углерод (С1), данный углерод
теряет связь с кислородом и соотвественно гликозидная связь разрушается и С1 связывается с кислородом аспартата 197. При этом рвется
водородная связь между Glu233 и кислородом гликозидной связи, при этом глутоминовая кислота в данной случае играет роль донора и отдает свой
протон. На этом первая стадия заканчивается. По сути первая реакция является реакцией нуклеофильного
замещения. Стадия дегликозилирования начинается с того, что одна из молекул воды приближается к аномерному углероду и его кислород-водородные
связи при этом начинают удлиняться, при этом одновременно с этим начинает разрушаться между С1 и аспартатом начинает разрушаться связь. Затем
данная молекула воды отдает свой протон Glu233, а вторая молекула воды отдает свой протон первой, восстанавливая ее. Теперь кислород гидроксильной
группы (которую образовала вторая вода) совершает нуклеофильную атаку на аномерный углерод, благодаря чему окончательно рарушается связь C1
c Asp197. Схема данной реакции представлена на рисунке 1. Данный механизм был описан в работах, которые мы прикрепили в разделе “Список литературы”[10][11]. Он интересен тем, что до этого было
множетсво работ которые также выдвигали теорию, что данная реакция основана на двух реакциях нуклеофильного замещения, но в них
говорилось только об участии одной молекулы воды. Тем не менее новое исследование показало, что это не так. Также до сих пор до конца
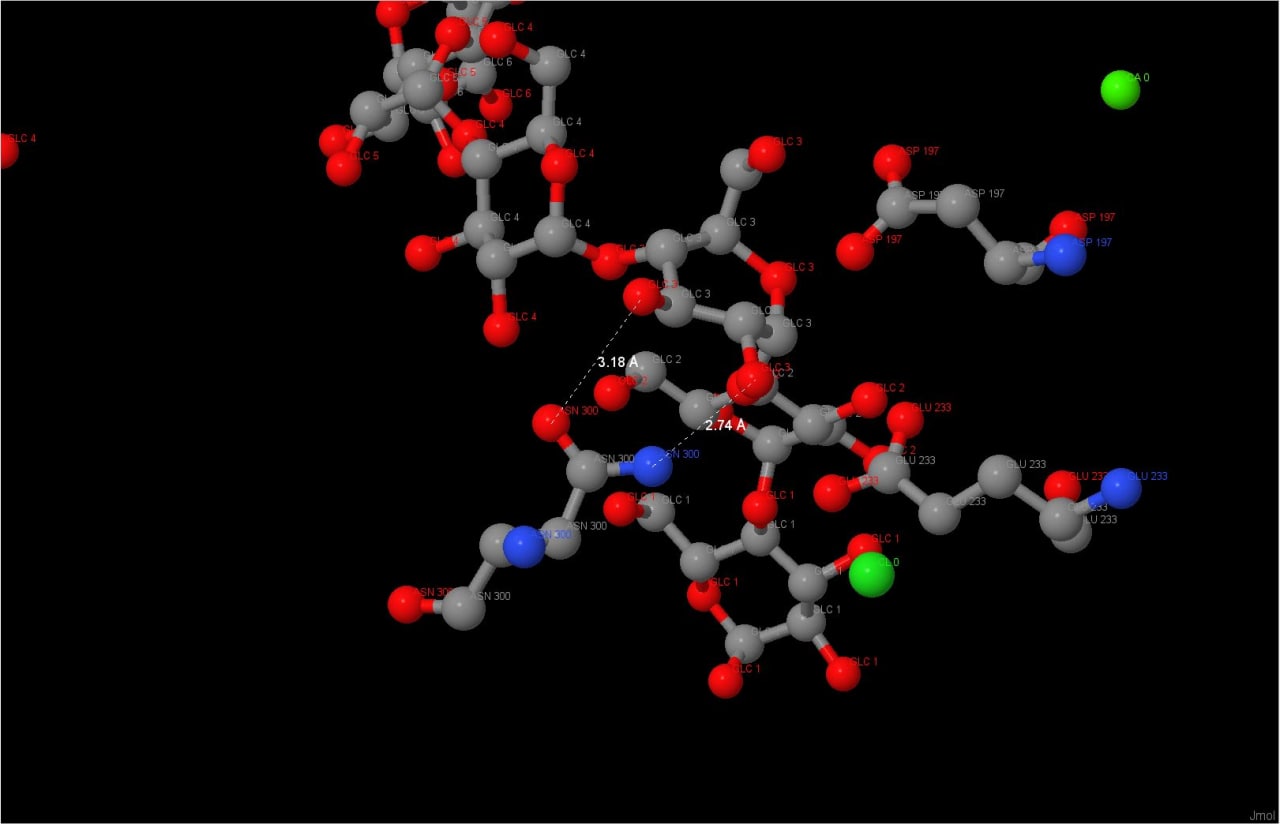
не ясна роль Asp300 в каталитической реакции. Тем не менее наш фермент содержит мутацию в данной позиции, а именно содержит остаток
аспаргина вместо аспаргиновой кислоты. Изучение аплета показало, что аспаргин находится на оптимальном
растоянии от гидроксильных групп кольца полисахарида, чтобы образовать с ними водородные связи (Рисунок 3), но
изучив статью на UniProt о нашем белке, можно заметить, что каталитическая активность сильно падает при
изменении Asp на Asn, поэтому роль Asp300 ещё предстоит изучить.
Рис.1 Механизм реакцииРис.2 Активный центрРис.3 Водородные связи
Водородные связи
Водородная связь — притяжение между ковалентно связанным атомом водорода и ковалетно
связанным электроотрицательный атомом, например O, N, F, Cl. Энергии водородной связи
достаточно, чтобы объединять молекулы в дилеры или полимеры. Посредствам водородной
связи так же образуются] вторичные, третичные и четвертичные структуры белков. Длина
водородной связи не превышает 3,5 ангстрем, иначе соединения между водородом и другим
электроотрицательный атомом не будет.[2]
Увидеть водородные связи можно выбрав скрипт "водородные связи" в аплете.
Имена атомов
Длина связи (Å)
N(MET 183) - O(ILE 179)
2.94
N(ASN 184) - O(ALA 180)
2.77
N(ILE 179) - O(VAL 175)
2.99
Таблица 1. Длины водородных связей
Солевые мостики
Солевые мостики — связь между положительно и отрицательно заряженными
аминокислотными радикалами, обусловленная возникновением электростатических
взаимодействий и водородных связей. Солевые мостики участвуют в формировании
третичной структуры белка.[3] Увидеть их можно выбрав скрипт "Солевые мостики" в аплете.
Для более удобного поиска солевый мостиков был использован ресурс http://pic.mbu.iisc.ernet.in/job.html.
На рисунке 1 изображен солевой мостик между GLU233 и ARG195, находящийся в непосредственной близости к активному
центру фермента.
Рис.1 Солевой мостик
Ковалентные связи
Дисульфидный мостик — ковалентная связь между двумя атомами серы, входящими
в состав цистеина, алифатической серосодержащей аминокислоты. Участвует в формировании
и стабилизации третичной структуры белка.[4] В молекуле находится несколько центров формирования
дисульфидной связи между молекулами цистеина. Нами, при помощи программы JMol, были выбраны центры,
находящиеся по краям молекулы. Затем, были замерены длины этих связей. Увидеть их можно выбрав скрипты
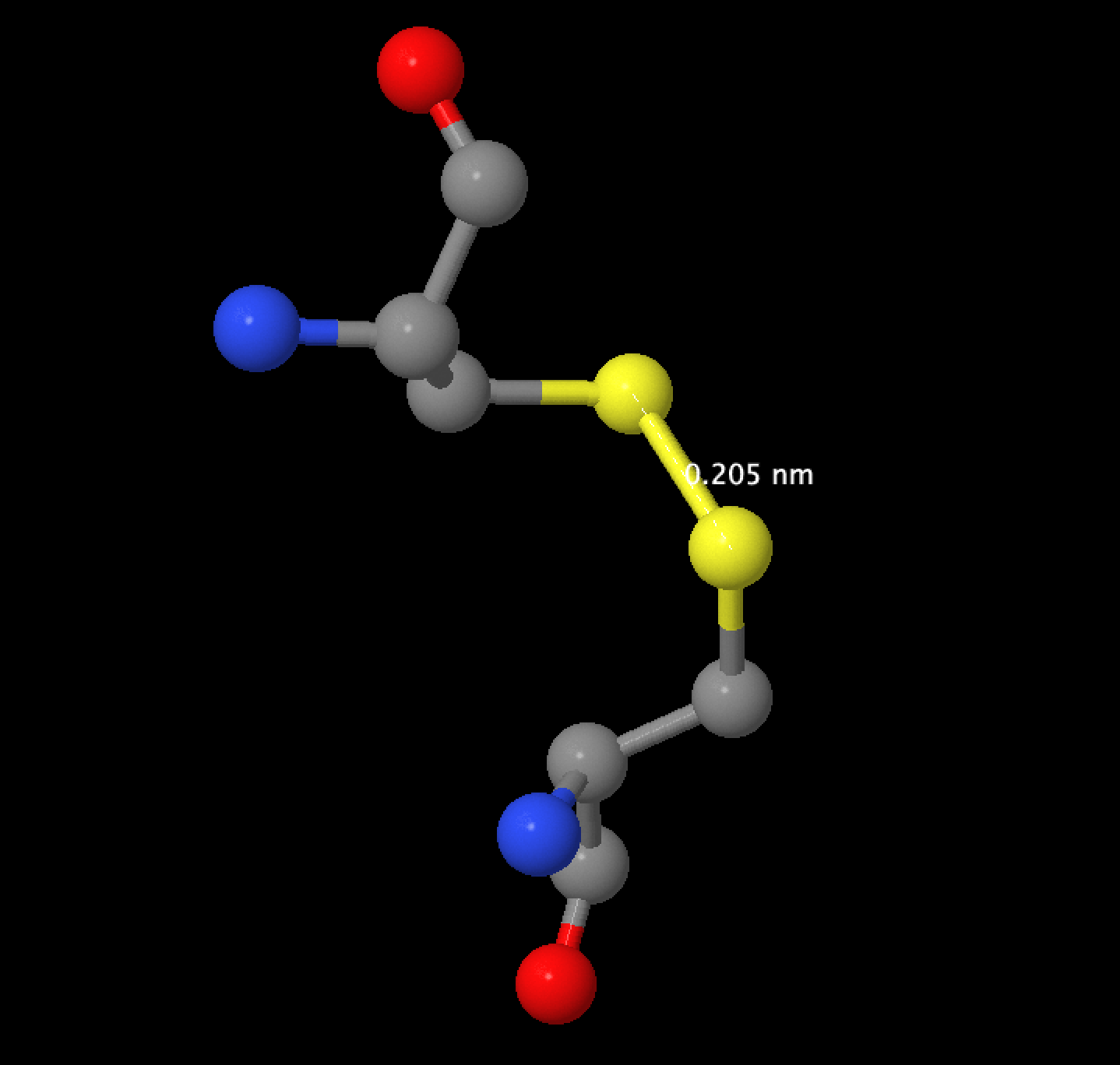
"Дисульфидный мостик 1" или "Дисульфидный мостик 2" в аплете. На рисунке 2 изображен дисульфидный мостик между CYS141 и CYS160, длина связи составляет 0,205 nm.
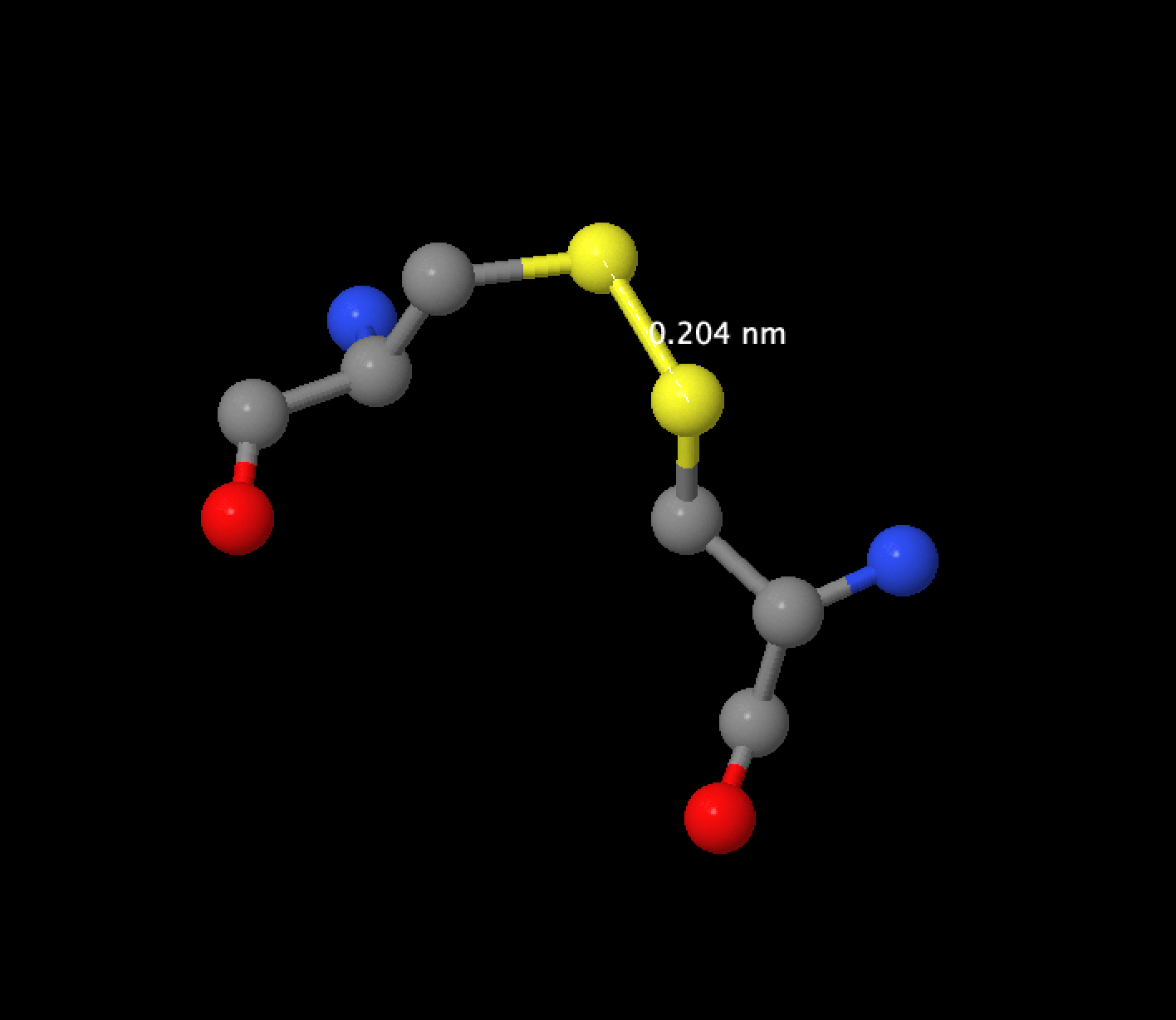
На рисунке 3 изображен дисульфидный мостик между CYS28 и CYS86, длина связи составляет 0,204 nm.
Рис.2 Дисульфидный мостик 1
Рис.3 Дисульфидный мостик 2
Стэкинг
Стекинг — тип нековалентного взаимодействия между ароматическими кольцами. В основе
стекинга находится межмолекулярное перекрывание p-орбиталей пи сопряженным систем.
Стекинг повышает стабильность молекулярной структуры.[5] Для облегчения поиска был
использован функционал сайта http://pic.mbu.iisc.ernet.in/job.html, который позволил
быстро обработать найденные структуры и замерить расстояние между двумя найденными
структурами. Увидеть это можно выбрав скрипт "Стэкинг 1" или "Стэкинг 2" в аплете.
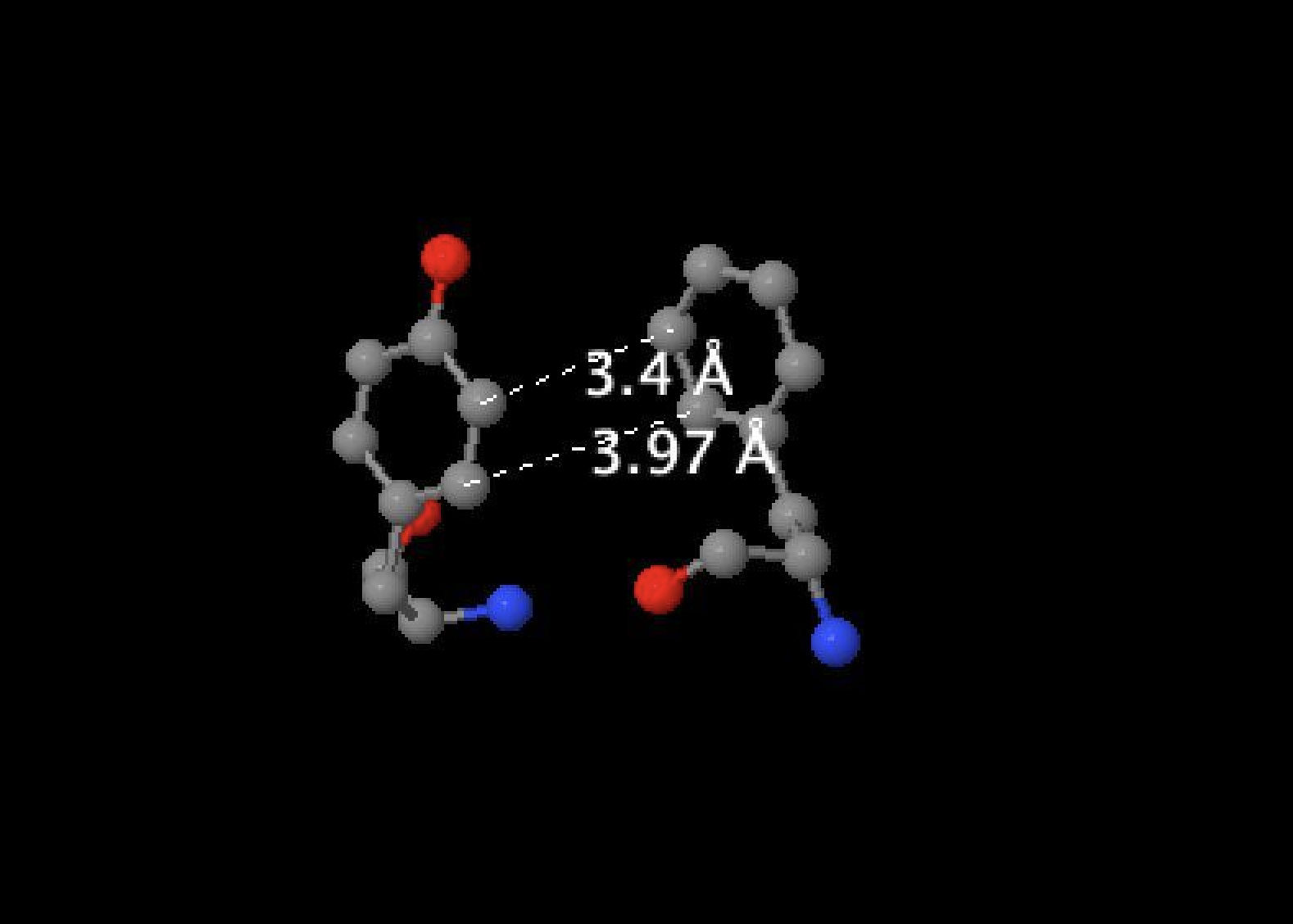
На рисунке 4 изображен стекинг между TRP 382 и TRP 19.
Рис.4 Стэкинг
Гидрофобное ядро
Плотность упаковки атомов в гидрофобном ядре.
Для исследования плотности упаковки атомов в гидрофобном ядре был выбран остаток фенилаланина [PHE]429:A.
В среднем, характерное расстояние между ядрами соседних не связанных ковалентно
атомами в белке не больше, чем 6.3Å ангстрем, т.е. в такое расстояние не поместится
молекула воды, поскольку расстояния между поверхностями атомов не больше 2.61Å, а диаметр атома кислорода составляет 2.8Å.[6]
Увидеть его можно выбрав скрипт "Гидрофобное ядро" в аплете. На рисунке 5 изображено гидрофобные ядра амилазы которые мы смогли найти
с помощью ресурса http://pic.mbu.iisc.ernet.in/job.html.
Рис.5 Гидрофобные ядра
Лиганды
Хлор
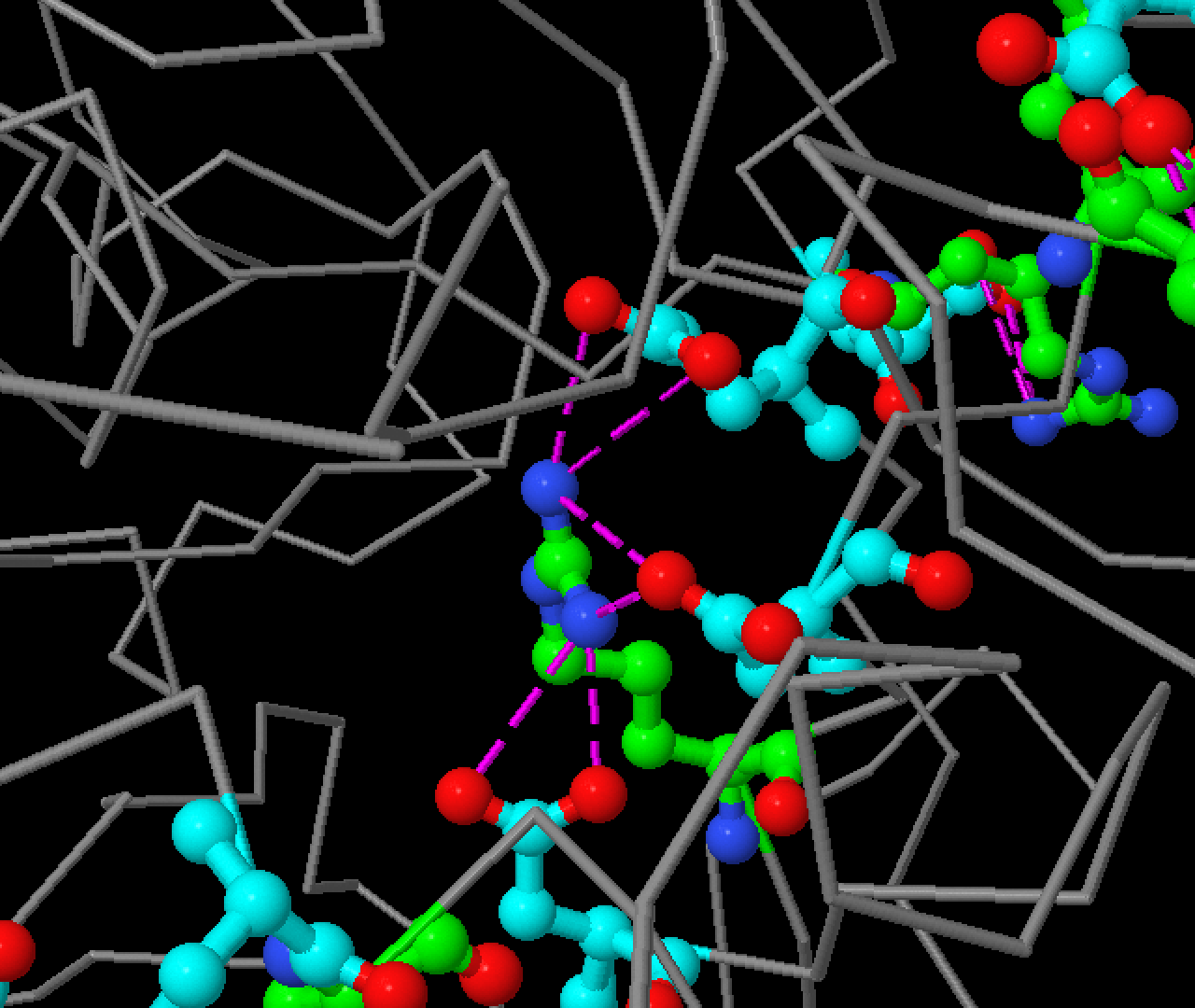
Хлорид ион находится очень близко к активному центру (ASN300, GLU233, ASP197) и расщелине связывания субстрата.
Он отвечает за активность и оптимальное значение pH белка.[7][8] Он был найден с помощью JMol.
Увидеть его можно выбрав скрипт "Хлорид ион" в аплете. На рисунке 6 изображен этот хлорид
ион и аминокислоты, входящие в активный центр амилазы.
Рис.6 Хлорид ион
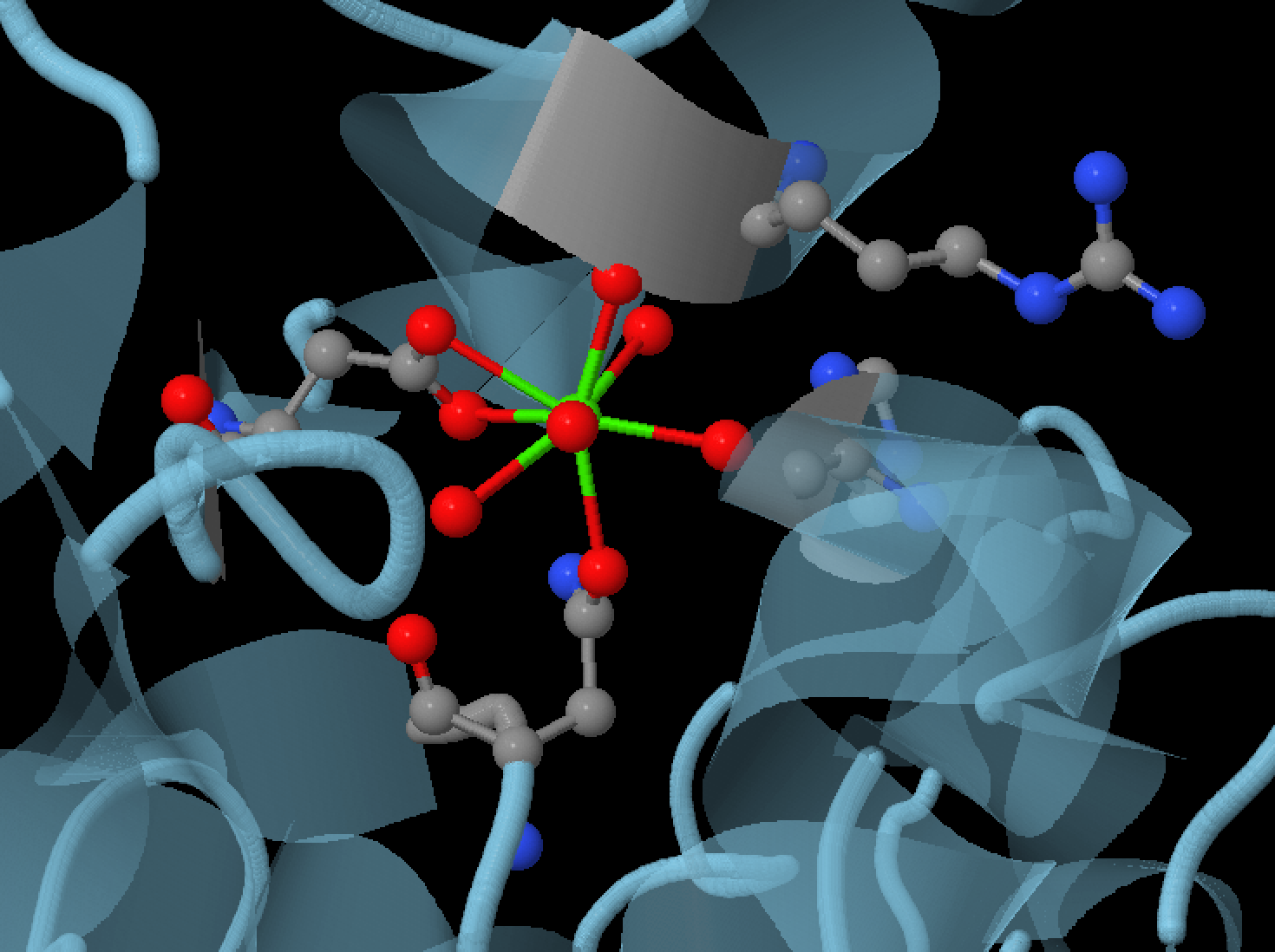
Кальций
Помимо хлора в соствв амилазы также входит и ион кальция. Он был найден тем же способом, что и хлорид ион. Ион кальция входящий в состав а амилазы обуславливает ее активность, он выступает в
роли акцептора электронов. Кальций очень прочно связан с амилазой и помогает ей сохранять ее структуру. Лишенный кальция фермент нестабилен и очень быстро теряет свою структуру.[9]
Увидеть его можно выбрав скрипт "Ион кальция в аплете".
На рисунке 7 изображен ион кальция, аминокислоты (ASN100, ARG1586, HIS201, ASP167), образующие с ним связи и молекулы воды,
которые также входят в комплекс кальция.
Рис.7 Ион кальция
Вклад авторов
Годованец Ольга - водородные связи, ковалентные связи, солевые мостики, стэкинг.
Кварталов Александр - низкомолекулярные лиганды, HTML страница.
Милейко Петр - гидрофобные ядра, работа с литературой.
Строгов Юрий - введение, основные понятия о связях и структурах.
6) Эмсли Дж. Элементы. (The Elements, 1991). Перевод с английского Е.А. Краснушкиной. Справочно-энциклопедическое издание.(Москва: Издательство «Мир». Редакция литературы по химии, 1993)
8) Активный центр: E. A. MacGregor, S. Janecek and B. Svensson (2001) Relationship of sequence and structure to specificity in the alpha-amylase family of enzymes. Biochimica et Biophysica Acta 1546, 1-20.
J. E. Nielsen and T. V. Borchert (2000) Protein engineering of bacterial alpha-amylases. Biochimica et Biophysica Acta 1543, 253-274.
10) Gaspar P. Pinto, Natercia F. Bra ́ s,́ Marta A.S. Perez, Pedro A. Fernandes, Nino Russo,
Maria J. Ramos, and Marirosa Toscano. Establishing the Catalytic Mechanism of Human Pancreatic
α‑Amylase with QM/MM Methods. Journal of Chemical Theory and Computation. Vol. 11, Issue 6, p. 2508-2516.
11) Gary D. Brayer, Gary Sidhu, Robert Maurus, Edwin H. Rydberg, Curtis Braun, Yili Wang, Nham T. Nguyen, Christopher M. Overall, and Stephen G. Withers.
Subsite Mapping of the Human Pancreatic α-Amylase Active Site through Structural, Kinetic, and Mutagenesis Techniques. Biochemistry. Vol. 39, Issue 16, p. 4778-4791.