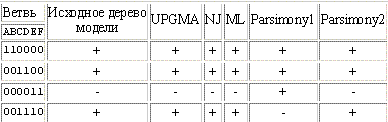
Мое исходное дерево:
Матрица эволюционных расстояний, определенных по методу Джукса - Кантора между последовательностями полученного выравнивания с помощью программы ednadist.
DXR_A 0.0000 0.4201 0.8908 0.8963 0.8638 1.3137 DXR_B 0.4201 0.0000 0.8586 0.8880 0.8404 1.3844 DXR_C 0.8908 0.8586 0.0000 0.2659 0.7837 1.4058 DXR_D 0.8963 0.8880 0.2659 0.0000 0.7861 1.4802 DXR_E 0.8638 0.8404 0.7837 0.7861 0.0000 1.4984 DXR_F 1.3137 1.3844 1.4058 1.4802 1.4984 0.0000
+-----------DXR_A
+-------------2
! +-----------DXR_B
+---------------4
! ! +------DXR_C
! ! +---------------1
--5 +--3 +------DXR_D
! !
! +----------------------DXR_E
!
+-----------------------------------------DXR_F
|
Это укорененное филогенетическое дерево построено по методу UGPMA. Данный алгоритм не является переборным, работает на основании матрицы расстояний, а не множественного выравнивания. Именно по этой причине данное построенное дерево является единственным полностью совпавшим по топологии с исходным. с помощью программы eneighbor Скобочная формула: (((DXR_A.:0.21005,DXR_B.:0.21005):0.22644,((DXR_C.:0.13295, DXR_D.:0.13295):0.25950,DXR_E.:0.39245):0.04404):0.27176, DXR_F.:0.70825); |
+------DXR_C
+--------------1
+-----3 +--------DXR_D
! !
! +-----------------------DXR_E
!
--4----------------------------------------------------------DXR_F
!
! +-----------DXR_A
+---------2
+------------DXR_B
|
Это дерево построено по методу ближайших соседей (Neighbor-Joining) с помощью той же программы eneighbor. Этот метод не строит дерево от листьев к предковым последовательностям, а разрешает изначально созданную небинарность. Логично, что созданное в ходе этого алгоритма дерево является неукорененным. Скобочная формула: (((DXR_C.:0.11899,DXR_D.:0.14691):0.25348,DXR_E.:0.39847):0.10226, DXR_F.:0.97599,(DXR_A.:0.20554,DXR_B.:0.21456):0.16301); |
+------DXR_B ! ! +------------------------------------------------DXR_F --1-----4 ! ! +-------------DXR_E ! +---3 ! ! +---DXR_D ! +--------2 ! +---DXR_C ! +------DXR_A |
Дерево, построенное по методу наибольшего правдоподобия (Maximum Likelihood) с помощью программы ednaml. Скобочная формула: (DXR_B.:0.23723,(DXR_F.:1.62582,(DXR_E.:0.46932, (DXR_D.:0.13961,DXR_C.:0.14325):0.29721):0.10824):0.19154, DXR_A.:0.22096); |
+--DXR_F
+-----5
! +--DXR_E
+--4
! ! +--DXR_D
+--2 +-----3
! ! +--DXR_C
--1 !
! +-----------DXR_B
!
+--------------DXR_A
|
Дерево, построенное по методу максимальной экономии (Parsimony) с помощью программы ednapars. Программа выдает на выходе два дерева. Это первое. Скобочная формула: ((((DXR_F.,DXR_E.),(DXR_D.,DXR_C.)),DXR_B.),DXR_A.)[0.5000]; |
+--------DXR_F
+--5
! ! +-----DXR_E
! +--4
+--2 ! +--DXR_D
! ! +--3
! ! +--DXR_C
--1 !
! +-----------DXR_B
!
+--------------DXR_A
|
А это второе дерево. Как видно из скобочной формулы этот метод не измеряет длины ветвей, является символьно- ориетированным. Скобочная формула: (((DXR_F.,(DXR_E.,(DXR_D.,DXR_C.))),DXR_B.),DXR_A.)[0.5000]; |