1. Выравнивание двух гомологичных последовательностей
По умолчанию программы используют для белков матрицу EBLOSUM62. Задается атрибутом -datafile
Штраф за открытие инделя - 10.0. В needle задается атрибутом -endopen, в water -gapopen
Штраф за удлинение инделя - 0.5. В needle задается атрибутом -endextend, в water -gapextend
Штраф за последний гэп по умолчанию отсутствует. В needle задается атрибутом -endweight
В отличие от программы needle, ориентированной на выравнивание последовательностей от первого до последнего знака, water выравнивает отдельные наиболее похожие участки последовательностей, вследствие чего могут быть потеряны некоторые буквы. В результате получившиеся выравнивания различаются между собой.
Сравнение основных параметров консервативности выравниваний, построенных в программах needle и water:
| программа | длина выравнивания | консервативные | функционально консервативные | позиции с гэпами | число инделей | ||
|---|---|---|---|---|---|---|---|
| число | процент | число | процент | ||||
| needle | 677 | 327 | 48.30% | 414 | 61,15% | 37 | 31 |
| water | 671 | 324 | 48.29% | 414 | 61,70% | 31 | 28 |
Water:

Needle:

Задание 2. Сравните параметры локального выравнивания пары гомологичных белков и пяти пар не гомологичных белков
При выравниваниии негомологичных последовательностей обнаружилось значительно меньшее количество консервативных позиций.
Характеристики выравниваний негомологичных последовательностей:
| идентификатор белка | длина выравнивания | консервативные | функционально консервативные | позиции с гэпами | ||
|---|---|---|---|---|---|---|
| число | процент | число | процент | |||
| A0A0X8D5E1 | 177 | 34 | 19,21% | 49 | 27,68% | 6 |
| E8YVR4 | 169 | 22 | 13,02% | 39 | 23,08% | 3 |
| A0A109QDP9 | 220 | 35 | 15,91% | 57 | 25,91% | 6 |
| A0A1B0VIM1 | 224 | 23 | 10,27% | 40 | 17,86% | 5 |
| A0A0B5QAX8 | 228 | 32 | 14,04% | 48 | 21,05% | 7 |
Заметно большее количество консервативных позиций в последнем выравнивании(гомологичных последовательностей):
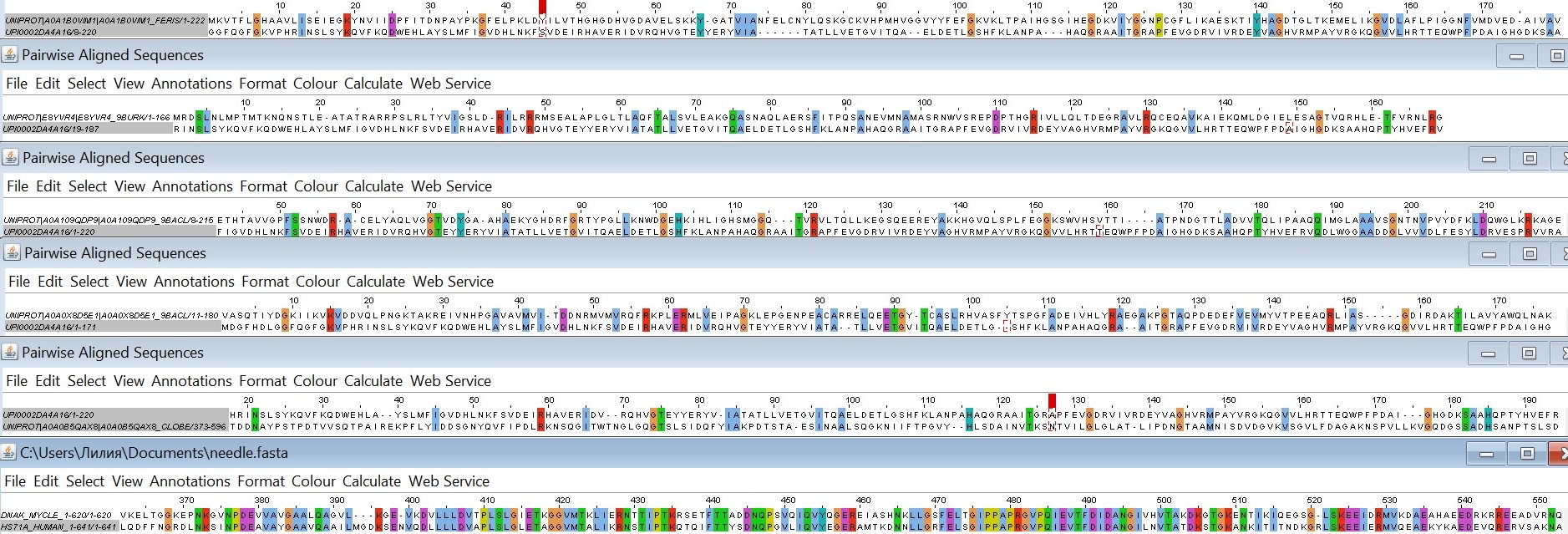
Сравнение выравниваний, построенных разными программами
Первые две последовательности вырезаны из множественного выравнивания. Третья и четвертая выравнены с помощью программы needle, пятая и шестая - water.
Выравнивания water и needle с параметрами по умолчанию различаются только тем, что water удалил 10 концевых позиций из первой последовательности.
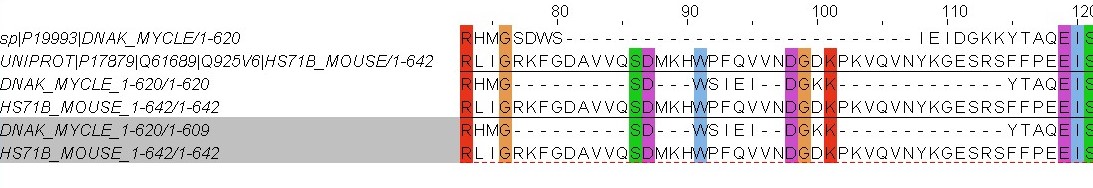
Парное выравнивание, полученное из множественного, совпадает с needle и water до 77 позиции, после в первой последовательности второго и третьего выравнивания стоит длинный индель. В результате серин с позиции 77 смещен на позицию 86 и выровнен с серином из второй последовательности. То же самое происходит со стоящей сразу за серином аспорагиновой кислотой.
В первом выравнивании между триптофаном(поз.79) иаспарагиновой кислотой(78) нет гэпа, и триптофан находится над фенилаланином второй последовательности. В needle и water между триптофаном(поз. 92) и аспарагиновой кислотой(поз. 87) стоит индель, и аспарагиновая кислота находится над аспарагиновой кислотой второй последовательности.
В первом выравнивании в первой последовательности аспарагиновая кислота и стоящий за ней глицин смещены на 111 и 112 позиции соответственно, и стоят над глутаминовой кислотой и серином второй последовательности. В выравниях water и needle они стоят на 98 и 99 позициях над идентичными аминокислотами второй последовательности.
Наиболее правдоподобным мне кажется первое выравнивание, так как оно построено с учетом положения соответсвующих аминокислот в других последовательностях. Чем больше последовательностей соответствуют выравниванию, тем больше вероятность правдоподобия гомологии аминокислот.