Авторы: студенты 1 курса факультета биоинженерии и биоинформатики Московского государственного университета им. М.В.Ломоносова Давитадзе Мария, Компаниец Мария, Неверов Александр, Холдина Дарья
1. Введение
В данной работе мы рассматриваем структуру папаина—цистеиновой эндопротеазы из млечного сока папайи (Carica papaya). Папаин представляет собой белок, состоящий из 212 аминоксилотных остатков, образующих одну полипептидную цепь (Otto & Schirmeister, 1997). Папаин выполняет функцию гидролиза белков в разнообразных участках цепи (практически нет ограничений по остаткам, см. далее). Мы работали с pdb-файлом структуры комплекса папина с лигандом E64c.
Идентификатор PDB: 1ppp.
Идентификатор белка в базе Uniprot: PRO_0000026407.
Код по международной классификации ферментов (EC): 3.4.22.2.
1.1 Катализ
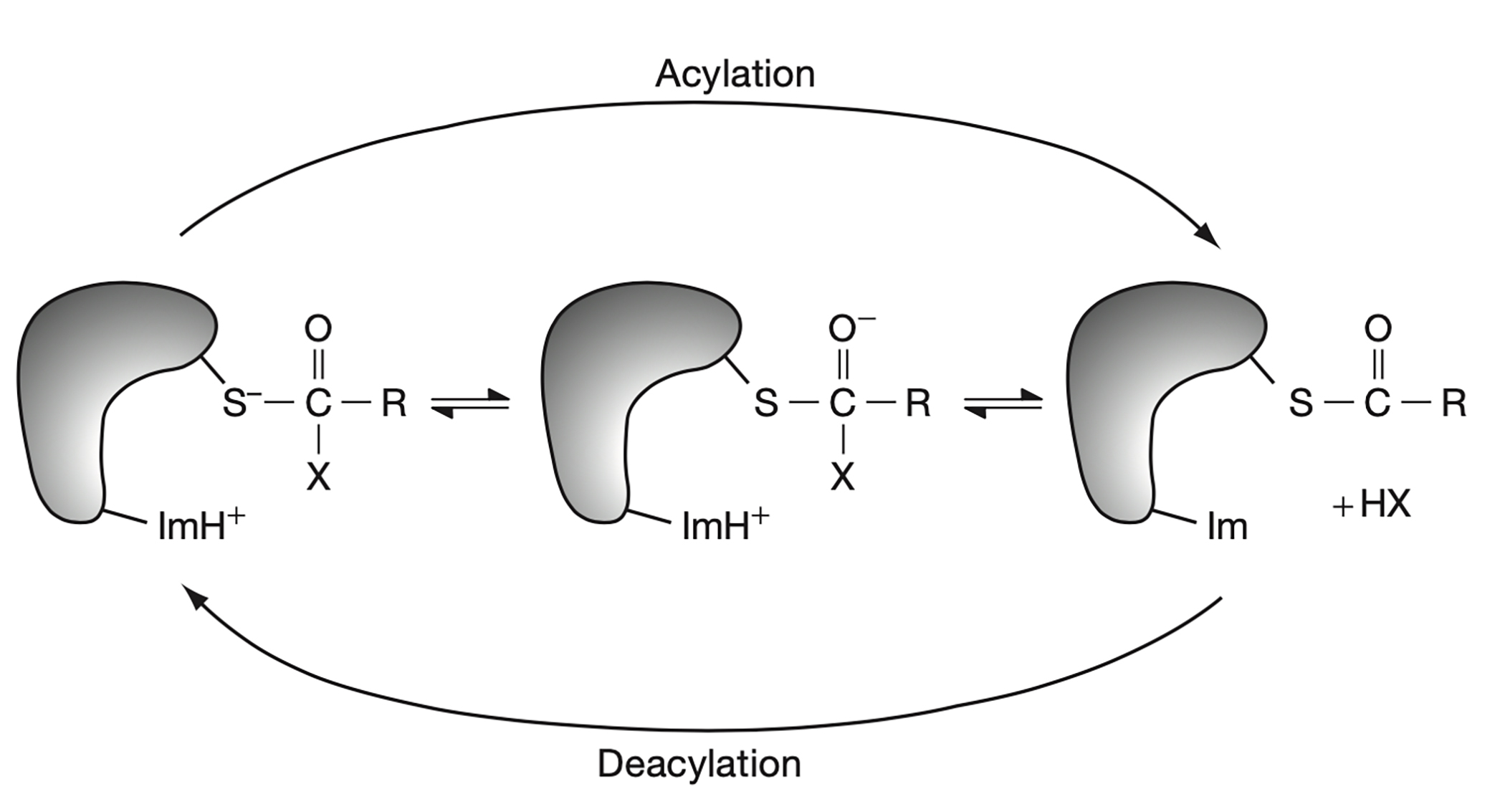
Активный центр папаина имеет характерную для цистеиновых протеаз структурную организацию и располагется в небольшом кармане в центральной части белка сверху. Непоредственно в реакции катализа участвуют два остатка: Cys25 и His159 (рис.1). В начале реакции тиольная группа на цистеине депротонируется с переходом протона на имидазол в составе гистидина в активном центре.
онированная сера атакует карбонил в составе субстрата, в результате чего образуется тетраэдрический интермедиат, стабилизирующийся остатками аспарагина и глутамина (Asn175, Gln19) (Novinec & Lenarčič, 2013). Далее с переносом протона с имидазола на гистидине на образовавшийся N-конец субстрата, последний высвобождается, а C-конец субстрата остаётся связанным тиоэфирной связью с остатком цистеина. Тиоэфирная связь затем гидролизуется, протон от молекулы воды, которая навешивается на углерод в составе карбонила, переходит на цистеин, восстанавливая тиольную группу, а C-конец высвобождается из активного центра (рис. 2). Остаток триптофана (Trp177) участвует в правильной работе каталитической пары (Gul et al., 2008). Механизм в целом описан по Buttle & Mort, 2013.
Рисунок 1. Изображение активного центра папаина с подписанными остатками, в первую очередь участвующими в катализе.
Рисунок 2. Схема реакции катализа, осуществляемой папаином (из Buttle & Mort, 2013).
1.2 Лиганды
В составе изученного нами комплекса помимо воды присутсвуют два лиганда: E64c (в структуре имеет код E6C, представлен одной молекулой) и метанол (MOH, представлен одной молекулой).
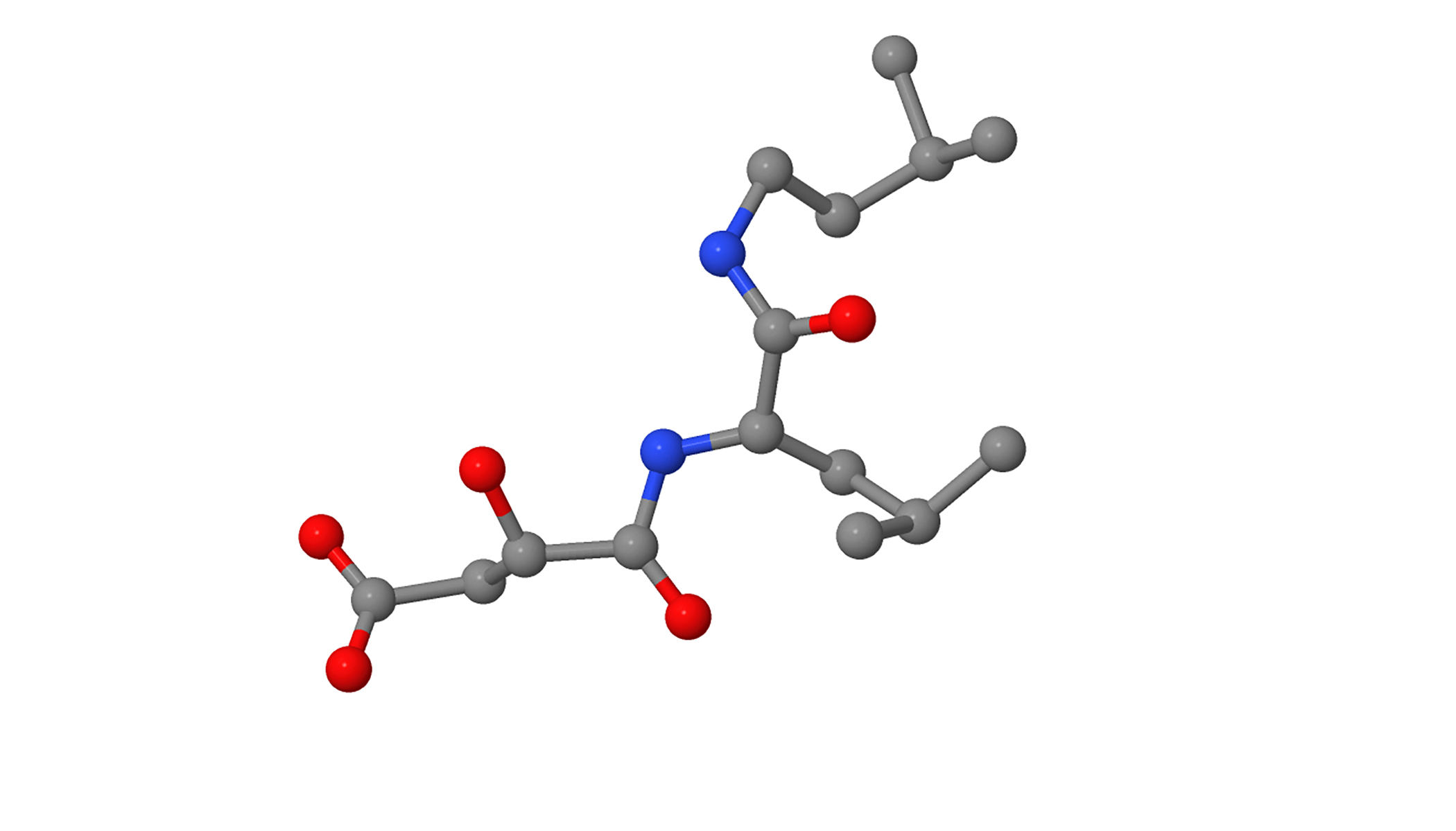
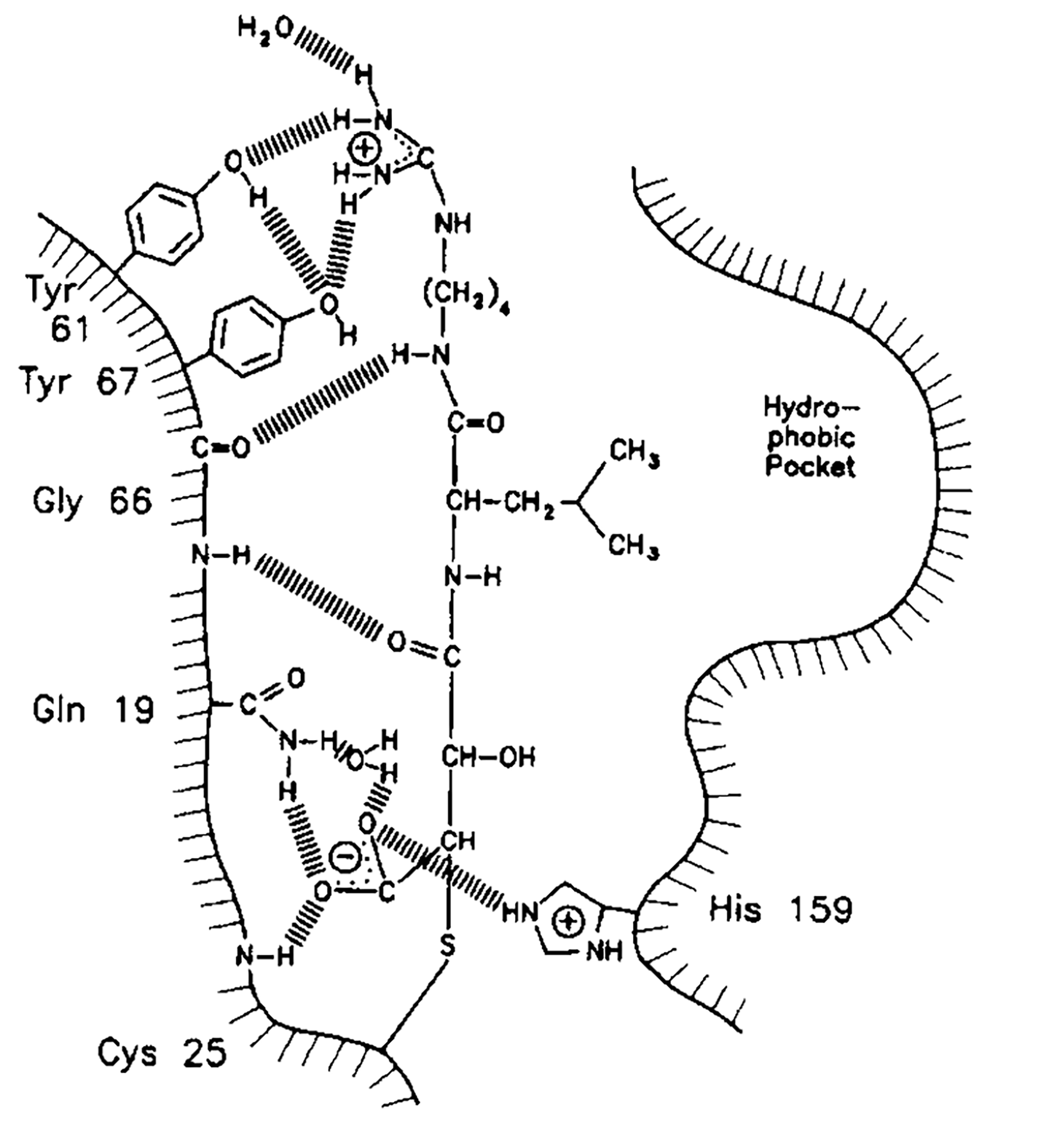
Лиганд E64c (рис. 3) является ковалентно связанным конкурентным ингибитором папаина (Otto & Schirmeister, 1997). Под названием E64c скрывается вещество N-[1-гидроксокарбоксиэтил-карбонил]-лейциламино-2-метилбутан, имеющее формулу C15H28N2O5 и молекулярную массу в 316.39 г/моль (идентификатор в базе данных PubChem: 5288147). Молекула этого ингибитора ковалентно связывается с цистеином в активном центре, образует водородные связи с боковыми радикалами аминокислот возле активного центра и закрывает остатком лейцина гидрофобный карман в активном центре (рис. 4). Таким образом, работа фермента становится невозможной.
Также в комплексе присутсвует молекула метанола (формула: CH4O, молярная масса: 32.042 г/моль, идентификатор в базе данных PubChem: 887). Упоминаний в литературе о функциях метанола в регуляции работы папаина мы не обнаружили. Возможно, он является примесью.
Рисунок 3. Изображение лиганда E64c, являющегося ингибитором папаина.
Рисунок 4. Механизм конкуретного ингибирования папаина лигандом E64c (из Otto & Schirmeister, 1997).
JMOL-апплет
Данный апплет последовательно показывает:
Лиганды вместе с белком
Активный центр белка
Водородные связи:
В альфа-спирали
Между бета-листами
Пи-водородные связи(2)
Два стэкинга
Расположение дисульфидных связей и пример дисульфидного мостика
Были рассмотрены водородные связи в альфа-спирали и бета-листах, а также π-водородные связи.
Водородной связью принято называть форму связи между электроотрицательным атомом (акцептор) и атомом водорода, прикрепленным ко второму, относительно электроотрицательному атому (донор). Участвующие электроотрицательные атомы обычно (но не обязательно) из первого ряда Периодической таблицы, т.е. N, O или F.
В белках водородные связи, образующиеся между карбонильной группой одного аминокислотного остатка и аминогруппой другого, играют важнейшую роль в поддержании вторичной структуры.
Как и любой тип связи, водородная связь имеет определенные структурные характеристики. Tак, для альфа-спирали идеальными считаются следующие значения длин связи и величин угла N-H-O: 2.99±0.14Å и 155±11°; для бета-листа: 2.91±0.14Å и 160±10° соответственно (Baker & Hubbard, 1984). Для нахождения водородных связей в нашем белке мы использовали команду calculate hydrogens, так как в нашей структуре отсутствовали атомы водорода, а затем calculate hbonds. По ходу выполнения программы обнаруживалось немало ошибок в виде, например, лишних водородных связей, поэтому в таблицу мы отобрали лишь некоторые из тех, которые не противоречат здравому смыслу. Как видно из таблицы 1, значения, полученные при анализе структуры папина, не сильно отклоняются от значений, приведённых в литературных источниках.
Таблица 1. Параметры водородных связей между остовными атомами во вторичной структуре
#
Имена атомов
Длина связи (Å)
Угол N-H-O (°)
Альфа-спирали
1
N(31VAL)-O(27ALA)
2.99
156.2
2
N(72LEU)-O(68PRO)
2.72
157.3
3
N(40ILE)-O(36GLY)
2.97
153.0
Среднее значение
2.89
155.5
Бета-тяжи
1
N(163ALA)-O(130VAL)
2.72
164.9
2
N(162ALA)-O(174LYS)
2.96
158.5
3
N(134LEU)-O(159HIS)
2.94
151.8
Среднее значение
2.87
158.4
π-водородная связь - это связь между донорной XH-группой и π-электронным облаком ароматического остатка. XH-группа располагается приблизительно над центром ароматического кольца, при этом на него указывает ось X-H связи. В π-водородных связях может наблюдаться довольно сильное смещение донора вбок относительно ароматического остатка, так что X-H связь может указывать как в центр ароматического кольца, так и в определенную C-C связь или даже на отдельный атом углерода. Встречается два основных типа геометрии π-водородных связей: первый, когда донорная группа перпендикулярна ароматическому кольцу, и второй, когда параллельна.
Энергия π-водородных связей ниже, чем у обычных O/N-HO водородных связей: в незаряженных системах она принимает значения от 2 до 4 ккал/моль, в заряженных же соответствующие показатели существенно выше. Расстояние между донором X и центром ароматического кольца обычно составляет от 3.2 до 3.8Å.
π-водородные связи, как и обычные водородные связи, играют роль в сворачивании белка и поддержании его структуры (Steiner & Koellner, 2001).
Для того, чтобы найти π-водородные связи, мы рассматривали ароматические аминокислотные остатки (Tyr, Phe, His, Trp), после чего проверяли, есть ли на подходящем расстоянии группа, содержащая свободный водород или частично положительный заряд для образования водородной связи с π-системой. При этом мы также смотрели, располагается ли эта группа параллельно или перпендикулярно ароматическому кольцу и нет ли рядом остатка, с которым эта группа могла бы образовывать обычную водородную связь.
Нам удалось найти несколько π-водородных связей, здесь мы покажем наиболее подходящие по параметрам. Мы измеряли длину связи по формуле нахождения медианы треугольника по трём сторонам.
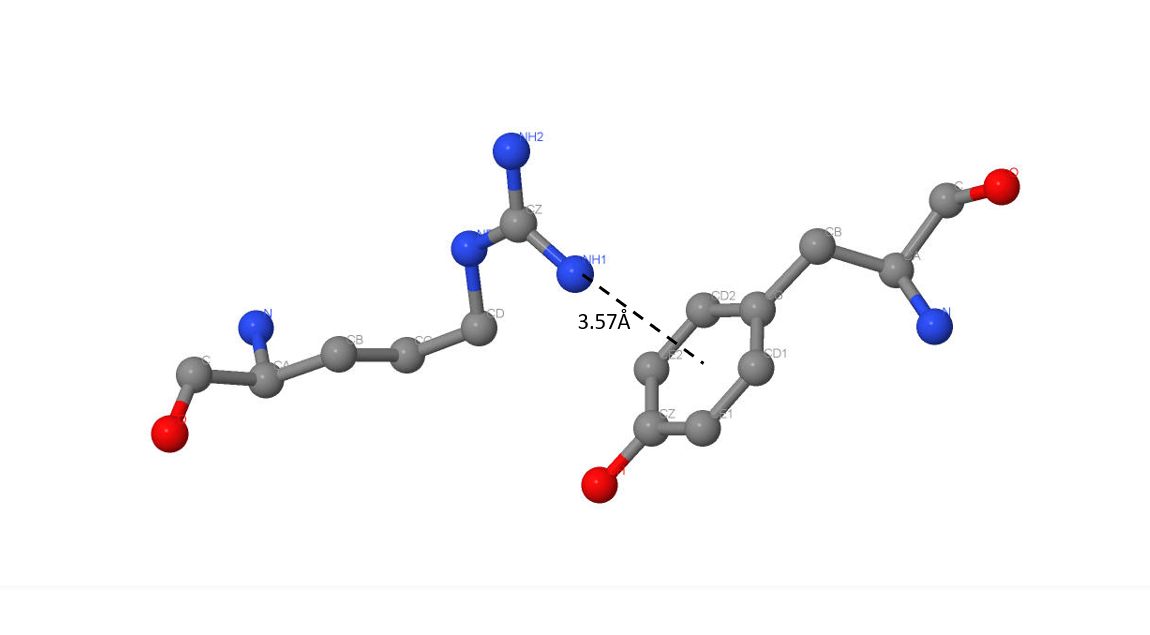
Длина π-водородной связи между [PHE]149 и [GLY]151 составляет 3,57Å (рис.5), а между [TYR]186 и [ARG]8 3,75Å от терминального азота радикала аргинина до центра кольца (рис.6).
Солевые мостики - pH-зависимые нековалентные связи, вызванные электростатическими взаимодействиями между противоположно заряженными остатками аминокислот. Их можно также рассматривать как особую форму водородных связей. Таким образом, солевые мостики могут возникать между отрицательно заряженными радикалами Asp, Glu, Tyr, Cys, а также терминальной карбоксильной группой и положительно заряженными остатками His, Lys, Arg и конечной аминогруппой (Bosshard et al, 2004). При этом расстояние между взаимодействующими группами не должно превышать 4.0Å (Kumar & Nussinov, 2002). Солевые мостики играют важную роль в поддержании структуры белка.
Мы искали солевые мостики с помощью программы Jmol. Мы использовали команды within (GROUP,(his or lys or arg) and within(4.0,asp or glu or tyr or cys)) и within (GROUP,(asp or glu or tyr or cys) and within(4.0,his or lys or arg)), чтобы выделить аминокислотные остатки, которые могли бы образовывать ионную пару, а затем искали остатки, заряженные группы которых располагались на подходящем расстоянии друг от друга.
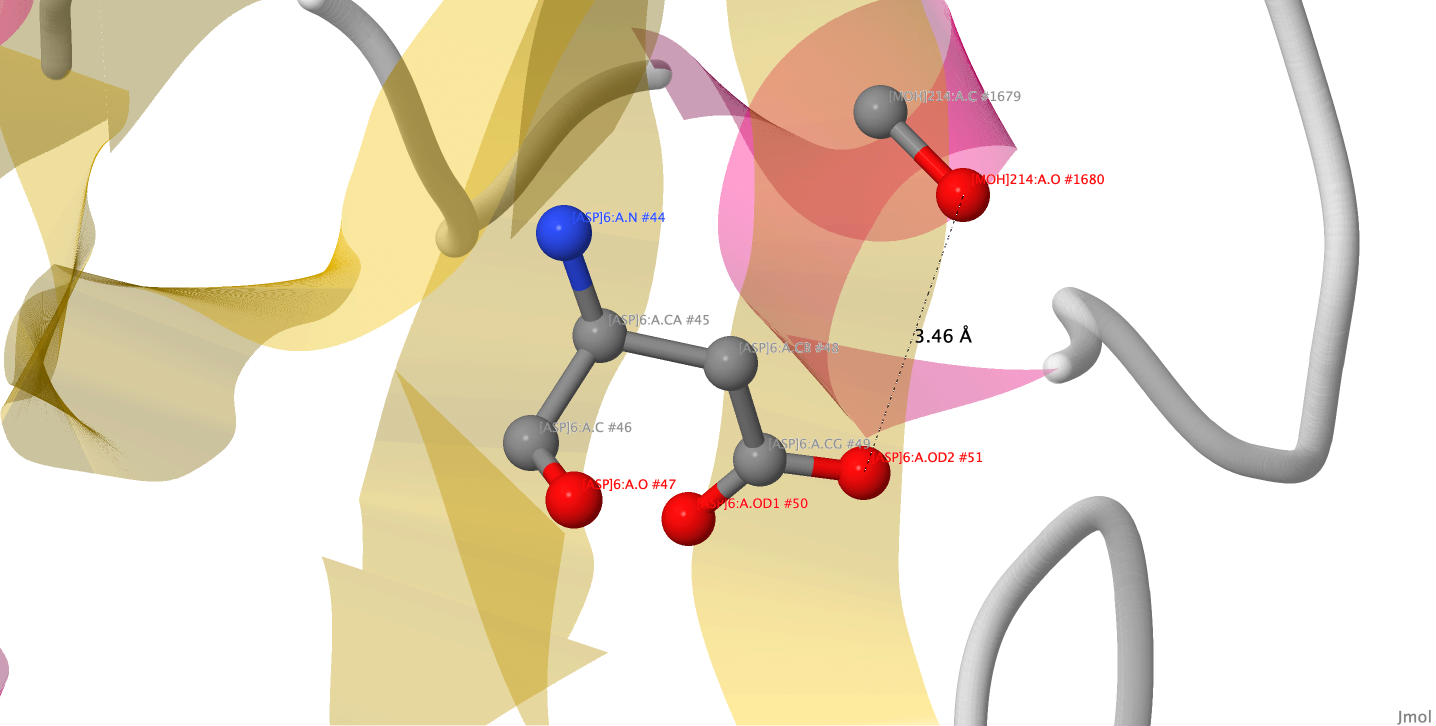
Рисунок 7. Солевой мостик между [ASP]6 и [ARG]8.
Рисунок 8. Солевые мостики [ASP]55-[ARG]96 и [GLU]89-[ARG]96.
c) Стэкинг
Стэкингом (или π-π-взаимодействием) называют нековалентные взаимодействия между ароматическими компонентами органических соединений. При этом происходит межмолекулярное перекрывание p-орбиталей в π-сопряженных системах (Martinez et al.,2012).
Считается, что для π-π-взаимодействия расстояние между центрами сопряженных систем должно быть не более 12Å (McGaughey et al.,1998). Для того, чтобы найти стэкинги, мы ввели команду restrict aromatic и вручную нашли аминокислотные остатки, находящиеся на наименьшем расстоянии друг от друга. Так, обнаружились как минимум два таких взаимодействия: между [TRP]181 и [TRP]177 (рис.9), а также между [TYR]82 и [TYR]48 (рис.10). Причём в первом из них угол между плоскостями очень близок к 90°, что наталкивает на предположение, что это Т-стэкинг. Во втором же случае мы наблюдаем параллельно смещенный тип стэкинга.
Рисунок 9. Стэкинг между [TRP]181 и [TRP]177.
Рисунок 10. Стэкинг между [TYR]82 и [TYR]48.
d) Ковалентные связи
Дисульфидные мостики и пептидная связь
К ковалентным относят два вида связей: пептидные связи и дисульфидный мостики.
Пептидная связь - вид ковалентной связи, которая возникает в результате взаимодействия α-аминогруппы одной аминокислоты и α-карбоксильной группы второй. Отвечает, в первую очередь, за формирование первичной структуры белка. Обычная длинна составляет 1.32Å (Berg JM et al., 2002). С некоторыми значениями длин пептидной связи в нашем белке можете ознакомиться в таблице 2.
Таблица 2. Длины пептидный связей
Имена атомов
Длина связи(Å)
N(67TYR)-C(66GLY)
1.31
N(112GLN)-C(111ARG)
1.32
N(19GLN)-C(18ASN)
1.32
Дисульфидный мостик - это вид ковалентной связи, возникающей между двумя атомами серы, входящими в состав двух остатков цистеина. Считается, что S-S-связь может оказывать значительное влияние на сворачивание белка и его стабильность (Matthias J. Feige et al., 2018).
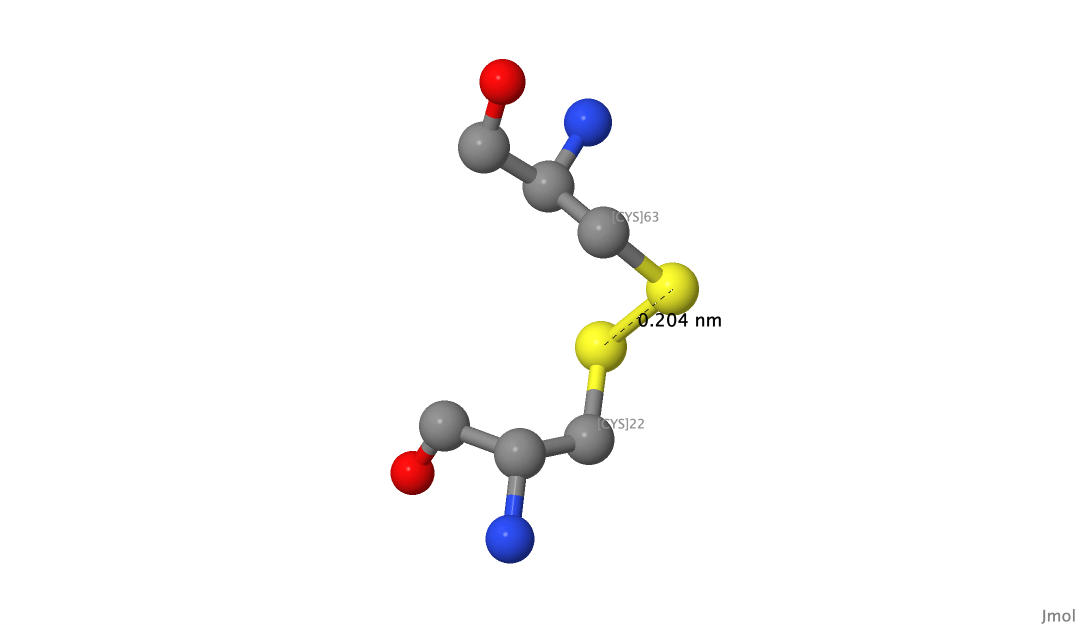
Чтобы найти S-S связи, мы ввели команду ssbonds on. Так, в структуре нашего белка было найдено 3 связи данного типа: [CYS]56-[CYS]95, [CYS]22-[CYS]63 и [CYS]153-[CYS]200. Их длины составляют 2.05Å, 2.04Å и 2.08Å соответсвенно, что соответствует нормальным значениям, которые в среднем составляют 2.05Å (Sun, Ming-An et al., 2017).
Рисунок 11. Расположение дисудьфидных связей.
Рисунок 12.Пример дисульфидного мостика между [CYS]22 и [CYS]63.
e) Гидрофобное ядро
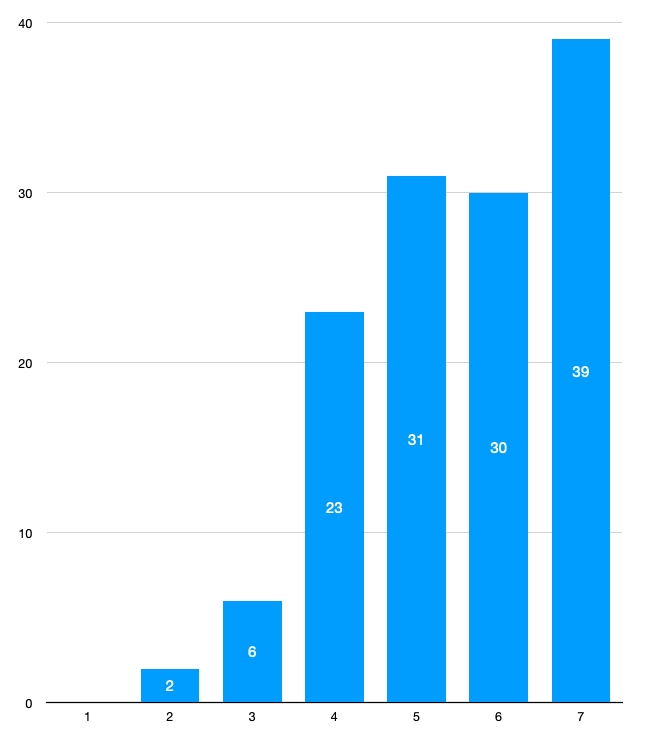
Для рассмотрения структуры упаковки гидрофобного ядра нами был выбран остаток фенилаланина под номером 141. Мы визуализировали атомы, располагающиеся на расстоянии от 1 до 7Å от выбранного остатка. Достаточное число атомов для почти сплошного покрытия остатка фенилаланина достигается на расстоянии 5-6Å от данного остатка (рис. 13). Также мы провели измерения расстояния между атомами в 25 парах ковалентно не связанных атомов на расстоянии 1-7Å от остатка фенилаланина. В среднем расстояние между атомами составило 4Å. На графике (рис.13), иллюстрирующем число атомов, нахоядящихся на конкретном расстоянии от фенилаланина, мы можем увидеть резкое увеличение числа атомов именно при достижении расстояния в 4Å.
Если брать среднее расстояние, то между атомами в упаковке гидрофобного ядра не поместится молекула воды (диаметр атома кислорода принимаем за 2.8Å (Emsley, 1993), то есть остается 1.2Å, чего не хватает, учитывая радиусы атомов в паре, между которыми могла бы поместиться вода).
Таким образом, упаковка в гидрофобном ядре является достаточно плотной. Между соседними атомами в большинстве случаев не может поместиться какой-то ещё из атомов, встречающихся в белках, кроме водорода.
Рисунок 13. Диаграмма распределения числа атомов в зависимости от расстояния от остатка фенилаланина 141 в гидрофобном ядре. По оси X отложено расстояние от остатка фенилалалнина в ангстремах, по оси Y и на столбцах отмечено число атомов, находящихся именно на этом расстоянии (атомы из N-того столбца не включаются в N+1). Видно резкое увеличение числа атомов окружения на расстоянии 4Å.
3. Лиганд-биомолекулярные контакты
Белок папаин связан с двумя лигандами: E64с и метанолом.
Лиганд E64c ковалентно связан с белком через цистеин. Также он образует водородные связи с остатками аспарагиновой кислоты и глутамина. Солевых мостиков и стэкинга лиганд с белком не формирует. Метанол же образует водородную связь с отрицательно заряженным остатком аспарагиновой кислоты.
Рисунок 14. Связь метанола с папаином.
Рисунок 15. Связь E64с с папаином.
Личный вкдлад каждого из участников
Мария Давитадзе осуществляла работу над описанием π-водородных связей, солевых мостиков, лиганд-биомолекулярных контактов.
Александр Неверов занимался написанием введения, а также работал над гидрофобными взаимодействиями.
Дарья Холдина работала над стэкингом, дисульфидными мостиками и водородными связями.
И, наконец, Мария Компаниец занималась версткой сайта, созданием аплета, модифицированием скриптов, а также координацией работы всей нашей команды.
Каждый этап работы сопровождался активным обсуждением всех участников.
Список литературы
D.J. Buttle, J.S. Mort (2013) Cysteine Proteases // Encyclopedia of Biological Chemistry. Elsevier Inc., P. 589-592.
E.N. Baker, R.E. Hubbard (1984) Hydrogen bonding in globular proteins // Prog Biophys Mol Biol., V. 44(2), P. 97-179.
G.B. McGaughey, M. Gagné, A.K. Rappé (1998) π-Stacking Interactions // Journal of Biological Chemistry, V. 273(25), P. 15458-15463.
H.-H. Otto and T. Schirmeister (1997) Cysteine Proteases and Their Inhibitors // Chemical Reviews, V. 97, P. 133-172.
H.R. Bosshard, D.N. Marti, I. Jelesarov (2004) Protein stabilization by salt bridges: concepts, experimental approaches and clarification of some misunderstandings // J. Mol. Recognit., V. 17, P. 116.
J.M. Berg, J.L. Tymoczko, L. Stryer (2002) Primary Structure: Amino Acids Are Linked by Peptide Bonds to Form Polypeptide Chains // Biochemistry. 5th edition New York: W.H. Freeman. Section 3.2.
M. Novinec, B. Lenarčič (2013) Papain-like peptidases: structure, function, and evolution // BioMol Concepts, V. 4, P. 287-308.
M.A. Sun, Y. Wang, Q. Zhang, Y. Xia, W. Ge, D. Guo (2017) Prediction of reversible disulfide based on features from local structural signatures // BMC Genomics, V. 18(1), P. 279.
M.J. Feige, I. Braakman, L.M. Hendershot (2018) Disulfide Bonds in Protein Folding and Stability (Chapter 1.1) // Oxidative Folding of Proteins: Basic Principles, Cellular Regulation and Engineering, P. 1-33.
Martinez, Chelsea, Iverson, Brent (2012) Rethinking the term "pi-stacking" // Chemical Science. 3 (7): 2191.
S. Gul, S. Hussain, M.P. Thomas, M. Resmini, C.S. Verma, E.W. Thomas, K. Brocklehurst (2008) Generation of nucleophilic character in the Cys25/His159 ion pair of papain involves Trp177 but not Asp158 // Biochemistry, V. 47.
S. Kumar, R. Nussinov (2002) Close-range electrostatic interactions in proteins // Chembiochem., V. 3, P. 604-17.
T. Steiner, G. Koellner (2001) Hydrogen bonds with π-acceptors in proteins: frequencies and role in stabilizing local 3D structures. Edited by R. Huber // Journal of Molecular Biology, V. 305(3).