1. Работа с KEGG PATHWAY
Общие сведения о метаболическом пути D-глутамина и D-глутамата
Все аминокислоты, в том числе глутамин и глутамат (рис. 1), входящие в состав молекул белков, являются L-стереоизомерами. В живой природе встречаются также и некоторые D-аминокислоты, но они никогда не входят в состав белков [1].
Примечательно, что неразличимые по своим обычным физическим и химическим свойствам энантиомеры зачастую резко отличаются друг от друга по физиологическому действию [2].
В данном биохимическом пути рассматриваются D-энантиомеры соответствующих аминокислот.
D-глутамин - это незаменимая аминокислота, которая в больших количествах присутствует в организме. Она является основным переносчиком азота в организме.
D-глутаминовая кислота не продуцируется эндогенно у высших млекопитающих и встречается в в клеточных стенках некоторых бактерий. Также D-глутамат является наиболее сильным природным ингибитором синтеза глутатиона, что может объяснить его локализацию в печени, поскольку D-глутамат может изменять окислительно- восстановительную стабильность [3].
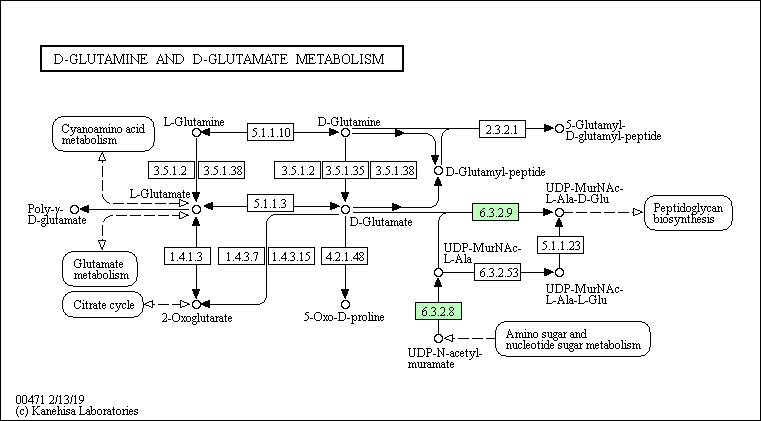
Согласно карте (рис. 2) D-энантиомеры глутамина и глутамата, которые являются начальными веществами в данной карте, превращаются в L-глутамат, который в последствии участвует в цианоаминокислотном метаболизме и метаболизме глутамата. Также метаболический путь D-глутамина и D-глутамата связан с синтезом пептидогликанов и циклом Кребса.
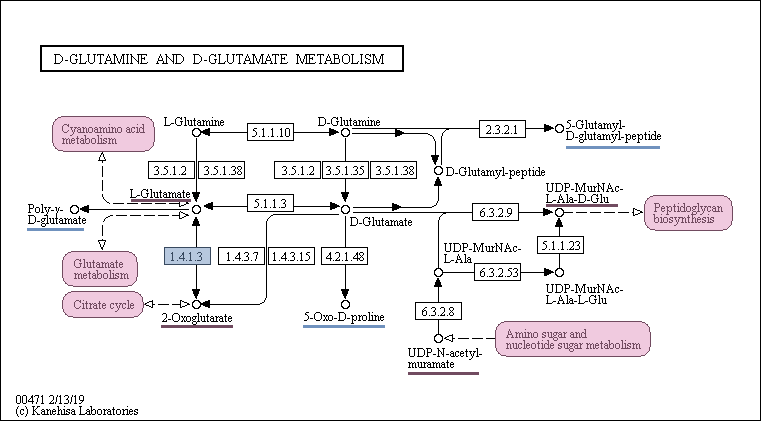
Метаболический путь D-глутамина и D-глутамата в разных доменах жизни
Из списка организмов, доступных для рассмотрения в БД KEGG, было выбрано три организма: Homo sapiens, Staphylococcus aureus MS4 и Methanobacterium paludis. Таксономия представлена в табл. 1.
| Вид | Homo sapiens | Staphylococcus aureus MS4 | Methanobacterium paludis |
| Вид | Человек разумный | Золотистый стафилококк | Нет названия на русском |
| Домен | Эукариоты | Бактерии | Археи |
| Тип | Chordata | Firmicutes | Euryarchaeota |
| Класс | Mammalia | Bacilli | Methanobacteria |
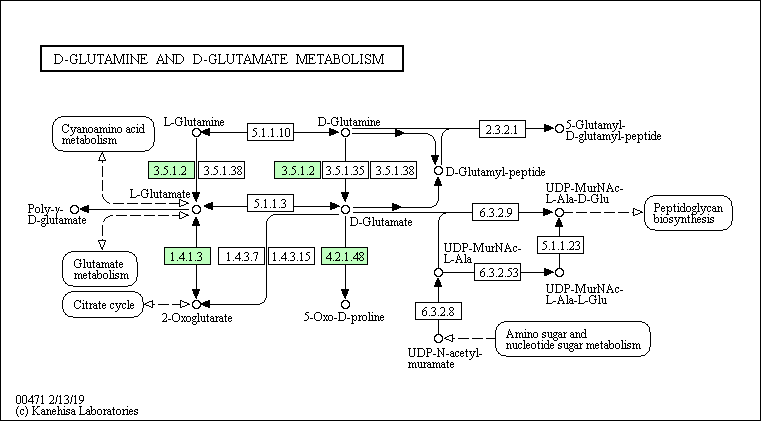
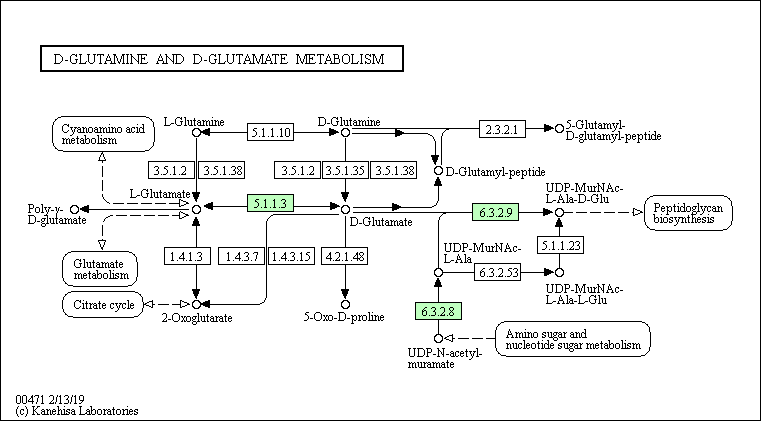
Ферменты, относимые к данному пути и присутствующие у каждого из выбранных организмов, отстутствуют. Тем не менее у представителя архей и представителя бактерий присутствуют общие ферменты - EC 6.3.2.8 (UDP-N-acetylmuramate---L-alanine ligase) и EC 6.3.2.9 (UDP-N-acetylmuramoyl-L-alanine ---D-glutamate ligase), являющиеся лигазами, которые катализируют образование C-N связи.
В каждом пути присутствуют короткие цепи последовательных реакций - максимум две реакции, которые не ведут от начальных для данного пути веществ к конечным, но есть цепочки превращений, ведущие в другие пути.
Так, Staphylococcus aureus MS4 и Methanobacterium paludis частично могут осуществлять данный путь, а именно тот, что ведёт к синтезу пептидогликанов (этот путь у человека полностью отсутствует, что можно объяснить тем, что у клеток человека отсутствует клеточная стенка, в состав которой входят пептидогликаны). В свою очередь, у человека есть ферменты, осуществляющие превращение D-глутамина в D-глутамат и L-глутамина в L-глутамат, но нет возможности превратить L-энантиомер в D - для глутамина и глутамата. Поэтому из начальных веществ (D-глутамин и D-глутамат) человек может получить только один конечный продукт - 5-Оксо-D-пролин.
Таким образом, выбранные организмы способны осуществлять данный путь лишь частично.
2. Работа с KEGG REACTION
Реакция дезаминирования L-глутамата в базе данных KEGG
Для метаболического пути D-глутамина и D-глутамата выбрана реакция, в которой принимает участие один из кофакторов окислительно-восстановительных реакций - никотинамидадениндинуклеотид (НАД). Идентификатор данной реакции: R00243. Данная реакция, а именно фермент, катализирующий реакцию, выделен синим цветом на рис. 2.
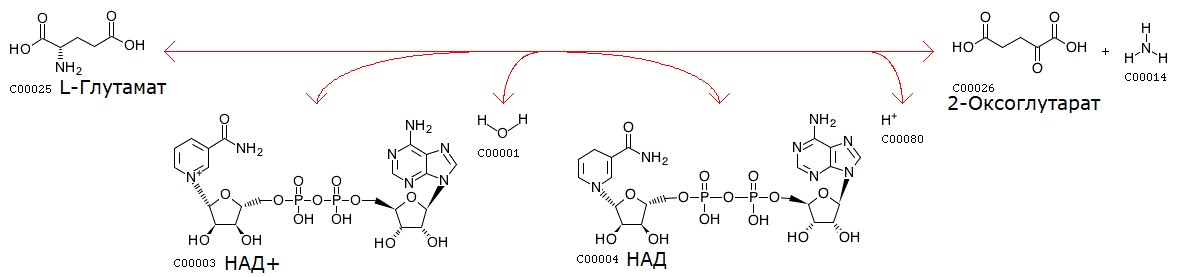
Схема реакции:
L-Glutamate + NAD+ + H2O <=> 2-Oxoglutarate + Ammonia + NADH + H+
C00025 + C00003 + C00001 <=> C00026 + C00014 + C00004 + C00080
3. Дополнительные задания
Выбранный мной путь для первого задания отсутствует, поэтому был выбран путь HIV.
мб потом сделаю и допишу этот пункт...4. Список литературы:
- Ленинджер А. Основы биохимии: В 3-х т. Т. 1. Пер. с англ.-М.: Мир, 1985.-367 с.
- Потапов В.М. Стереохимия; Учеб. пособие для вузов.— 2-е изд., перераб. и доп.— М.: Химия, 1988. —464 с.: ил.
- Raj, D., Langford, M., Krueger, S., Shelton, M. & Welbourne, T. Regulatory responses to an oral d - glutamate load: formation of d -pyrrolidone carboxylic acid in humans. Am. J. Physiol. Metab. 280, E214–E220 (2001).