Практикум 3
Задание 1. Предсказание вторичной структуры заданной тРНК
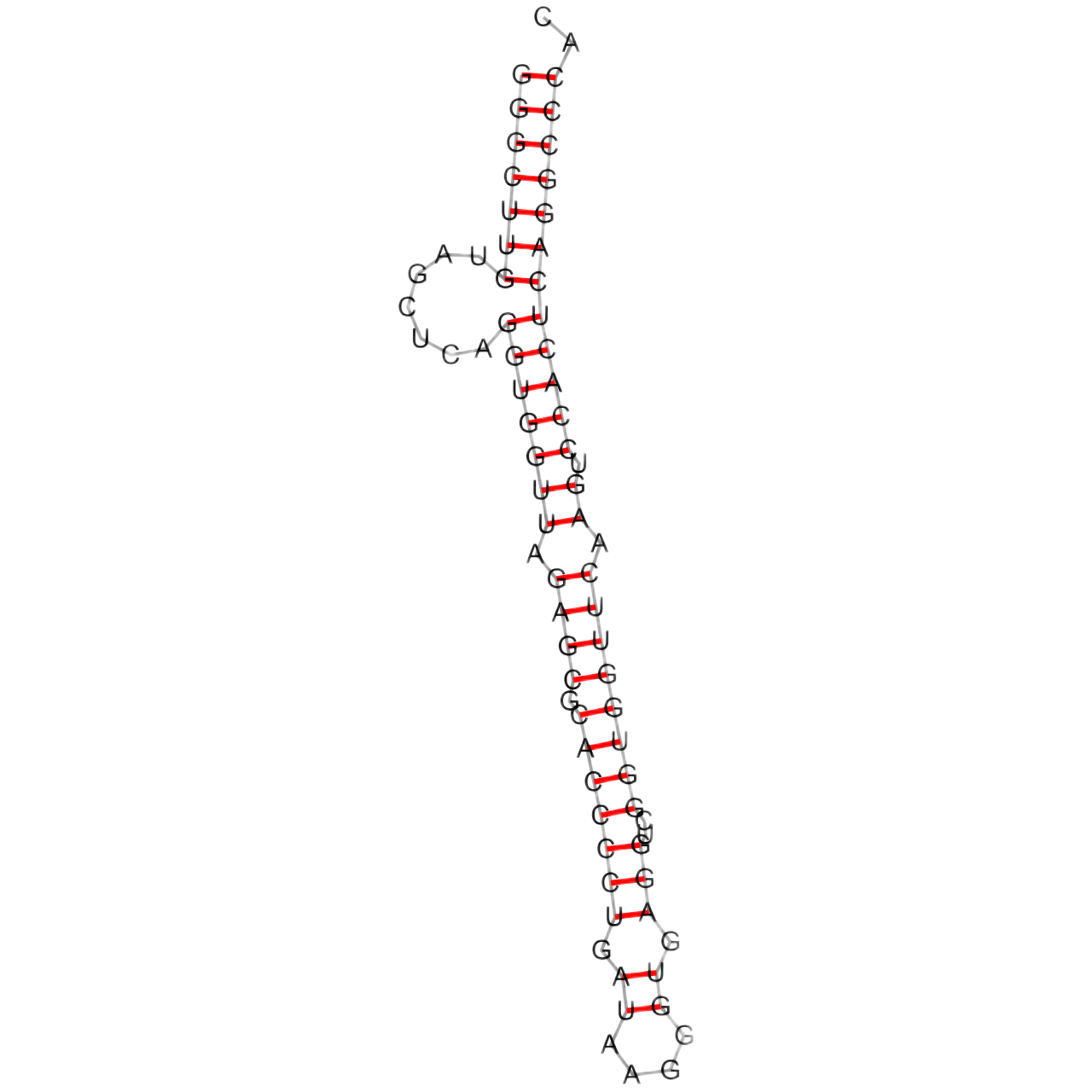
При помощи программы einverted из пакета EMBOSS были найдены возможные комплементарные участки в последовательности исследуемой молекулы (1ffy), а также по алгоритму Зукера была предсказана вторичная структура тРНК с помощью ViennaRNA (Рис.1).
Затем результаты были сравнены с описанием, полученным при помощи программы find_pair (Табл.1).
| Участок структуры | Позиции по find_pair | Результаты einverted | Результаты предсказания по алгоритму Зукера |
|---|---|---|---|
| Акцепторный стебель | 5'-1...7-3' 5'-66...72-3' | 2/7 | 5'-1...7-3' 5'-66...72-3' (7/7) |
| D-стебель | 5'-10...13-3' 5'-22...25-3' | 0/4 | 0/4 |
| T-стебель | 5'-49...53-3' 5'-61...65-3' | 0/5 | 0/5 |
| Антикодоновый стебель | 5'-38...44-3' 5'-26...32-3' | 2/7 | 0/7 |
| Общее число канонических пар нуклеотидов | 23 | 14 | 22 |