PubMed
Историю запросов и результатов поиска белка RB в PubMed
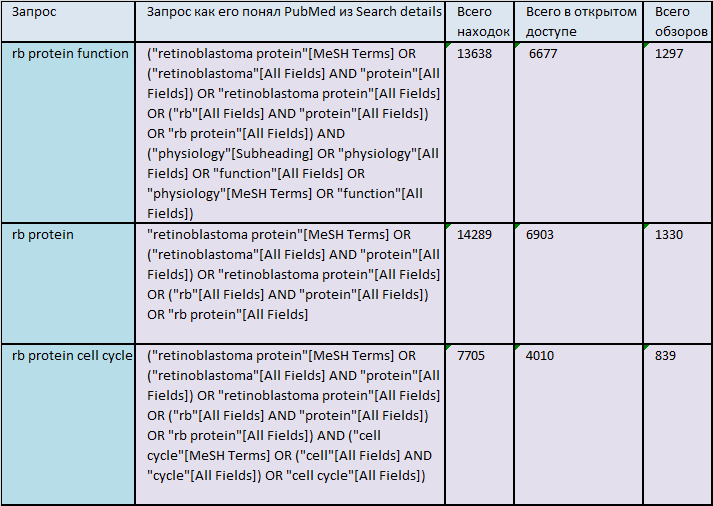
Персональная коллекция публикаций о белке retinoblastoma rotein(RB)
В ходе поиска публикаций о белке RB в Pubmed была создана коллекция, содержущая статьи описавающие функции белка в клетке. Ссылка на неё представлена ниже:
Коллекция публикаций о белке функциях белка в клетке retinoblastoma rotein.
Краткое описание статьи: "Rb protein down-regulates the stress-activated signals through inhibiting c-Jun N-terminal kinase/stress-activated protein kinase." Shim J, Park HS, Kim MJ, Park J, Park E, Cho SG, Eom SJ, Lee HW, Joe CO, Choi EJ.
Rb-белка регулирует снижение стресс-активируемых сигналов путем ингибирования C-Jun N-концевой киназы / стресс-активируемой протеинкиназы.
Введение
Rb белок является продуктом работы гена ретинобластомой восприимчивости. Во многих типах опухолей человека были обнаружены потереи функции Rb. Rb играет важную роль в регуляции клеточной пролиферации, дифференцировки, а также клеточного старения и апоптоза. Эксперемент показывает, что Rb может физически взаимодействовать с C-Jun NH 2-концевой киназой/стресс-активированной протеинкиназой (JNK/SAPK), тем самым препятствавать внутриклеточным сигналам посредствам JNK/SAPK. Ген ретинобластомной восприимчивости, продуктом котрого является белок ретинобластомы (Rb), негативный регулятор роста клеток, путем подавления перехода от G1 до S-фазе клеточного цикла. Rb действуют как ингибиторы роста клеток и опухолевых супрессоров, взаимодействую с фактором транскрипции E2F и другими клеточными белками. Например, нефосфорилированная или гипофосфорилированная форма Rb, путем связывания E2F, подавляет экспрессию E2F-регулируемые гены, необходимого для синтеза ДНК, что приводит к остановку клеточного цикла. Rb связываться с другими факторами транскрипции: Elf-1, MyoD, PU.1, UBF, с-Мус и ATF-2; а также другими регуляторными белками, такими как гистондеацетилаза, MDM2, и с-Abl ( 16-20 ). Возможность ретинобластомы взаимодействовать с различными белками что говорит о её многофункциональности и способности регулировать не только клеточный цикл, но и клеточную активность. Недавние исследования показали, что Rb может регулировать апоптоз. Кроме того, во время апоптоза Rb может быть расщеплена, на СООН-конце, по каспазе(caspases) и ослабляеть антиапоптозным функции активной форме Rb. Молекулярный механизм по борьбе со смертным влиянием Rb остается неясным. Даное исследовании демонстрируют новые функции Rb, который подавляют JNK/SAPK сигнальные пути через ингибирования деятельности JNK/SAPK. Ингибирование JNK/SAPK активности может быть важным механизмом, котрый негативно регулирует Rb, вызывая напряжение активированных клеточных событий, в том числе гибель клеток.
Экспериментальные методики
Клетки НЕК293 Saos-2 обычно поддерживали в среде Eagle's, модифицированной Dulbecco's, c добавкой 10% фетальной бычьей сыворотки. Клетки трансфицировались соответствующими векторами экспрессии посредством LipofectAMINE (Life Technologies, Inc.). После 40-48 ч трансфекции, клетки подвергали воздействию ультрафиолетового света (60 Дж/м2).
Ядра были получены из культивируемых клеткок НЕК293 описанным выше способом и осаждены либо с полученными от нескольких клонов anti-Rb антителами, либо устойчивыми к негативным эффектам IgG. Иммунопреципитаты подвергали электрофорезу на 12% SDS-полиакриламидном геле, иммуноблоттинг моноклональных антител (Pharmingen) исследовали на мыши anti-JNK1/SAPKγ и визуализировали с помощью метода усиленной хемилюминесценции (Amersham Pharmacia Biotech).
Культивируемые клетки были собраны и лизированны в лизисном буфере. Растворимые фракции подвергли конденсации с соответствующими антителами и полученные имунные гранулы химически проанализировали на индуцированную протеинкиназу. Фосфорилированные белки были разделены электрофорезом в 10% SDS-полиакриламидном геле и помечены с помощью метода автоматической рентгенографии и измерения плотнотсти. GST-С-Jun( 1-135) или GST-ATF2 (1-109) были использованы в качестве субстрата для JNK, GST-ATF2 (1-109)- для р38,белок миозин - для ERK. GST-SEK1 (K129R) использовали в качестве субстрата для анализа MEKK1. Мышиные моноклональные анти-Flag М2 (Kodak), анти-HA (Roche Molecular Biochemicals) или анти-JNK1 антитела были использованы в качестве индикаторных.
HEK293-клетки трансфицировали с помощью вектора pEBG, экспрессирующего GST-SEK1. Трансфицированные клетки лизировали, и солюбилизировали с 1% Triton Х-100. Солюбилизированную фракцию нанесли на глутатион-агарозных и GST-SEK1 элюировали из смолы, после чего анализировали на активность с использованием SEK1 гексагистидиновой-меченый p38, в качестве субстратного белка.
SAPKβ и ERK1 были в пробирке переобразованы с ретикулоцитами кролика системой лизаты (Promega) с использованием [35S]-метионина. Для измерения преобразованых in vitro инкубировали в течение 2 ч при 4 ° С с указанным слитых белков GST иммобилизованных на глутатион-сефарозы в связывающем буфере и гранулы промывают широко промывочным буфером. 35 S-меченые белки элюируют из шариков, разделенных электрофорезом на 10% ДСН-полиакриламидном геле и визуализировали с помощью авторадиографии.
SHEK293-клетки трансфицировали с помощью вектора pEBG, экспрессирующего GST-SEK1. Трансфицированные клетки лизировали, и растворили в 1% Triton Х-100. Растворимую фракцию нанесли на glutathione-agarose,GST-SEK1 элюировали из смолы, после чего проанализировали на активность SEK1 с использованием hexahistidine-tagged p38 в качестве субстратного белка.
Результаты
Для того чтобы узнать о возможности Rb воздействать на функцию пути JNK/SAPK, было исследовано действие Rb на JNK активность в культуре клеток. Клетки НЕК293, трансфицировали с вектором, экспрессирующим SAPKβ-флаг*, или JNK1-HA, или провели это без вектора, экспрессирующего полностью белок Rb. SAPKβ и JNK1 идентичны JNK3 и SAPKγ соответственно. JNK активности стимулируется УФ-облучение трансфицированных клеток (рис. 1 ).
Рис. 1. Rb ингибирует JNK / SAPK в интактных клетках Rb ингибирует JNK / SAPK, послеле УФ облучения в НЕК293. HEK293 клетки нетрансфецированных или трансфицированные кДНК конструкции для SAPKβ-флаг или JNK1-HA и Rb, как указано. После 48 ч трансфекции клетки облучают УФ-излучением (60 Дж/м2) и инкубировали еще в течение 1 часа. Ферментативных активностей эктопически выразил JNK в трансфицированные клетки измеряется иммунокомплексными анализа киназы.
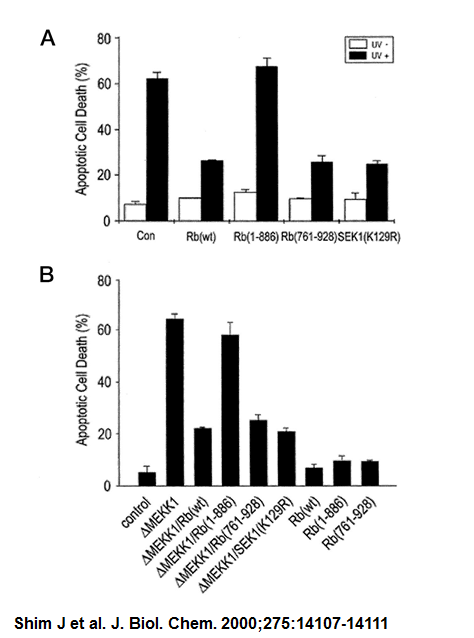
Результаты в этого исследования заключается в том, чтобы предпложить новую функцию RB: RB ослабление функций JNK/SAPK пути. Из в ходе эксперемента было показано, что Rb может подавлять стресс-активированных сигналы, идутущие по этому самому пути(пути JNK). Также вызывает интерес то, что Rb может расщепляется на каспазы, и тем самым противодействовать апоптозу клетки. Такое расщипление проявляется потерей функцией анти-клеточной гибели. Расщепление происходит в СООН-концевой области между Asp 886 и Gly 887, генерируя Rb-белк, который не хватает 42 аминокислот. Каспаза, которая образовалась после расщепления, еще может взаимодействовать с Rb-связывающими белками, такими как E2F или циклин D также эффективно, как полноценый белок Rb. Но есть отличие между формами есть отличие, расщепленная не может препятствовать JNK пути. Это может означать, что JNK может быть критической целью Rb для того, чтобы подавить стресс-индуцированное гибели клеток. Таким образом, ингибирование JNK/SAPK может быть новая функция Rb, что негативно регулирует активированные сигнальные каскады.
-флаг* - надстройка на молекулу, за которую её можно увести(опазнать и оттянуть)